

AAV-mediated, optogenetic ablation of Müller Glia leads to structural and functional changes in the mouse retina
- PMID: 24086689
- PMCID: PMC3785414
- DOI: 10.1371/journal.pone.0076075
AAV-mediated, optogenetic ablation of Müller Glia leads to structural and functional changes in the mouse retina
Abstract
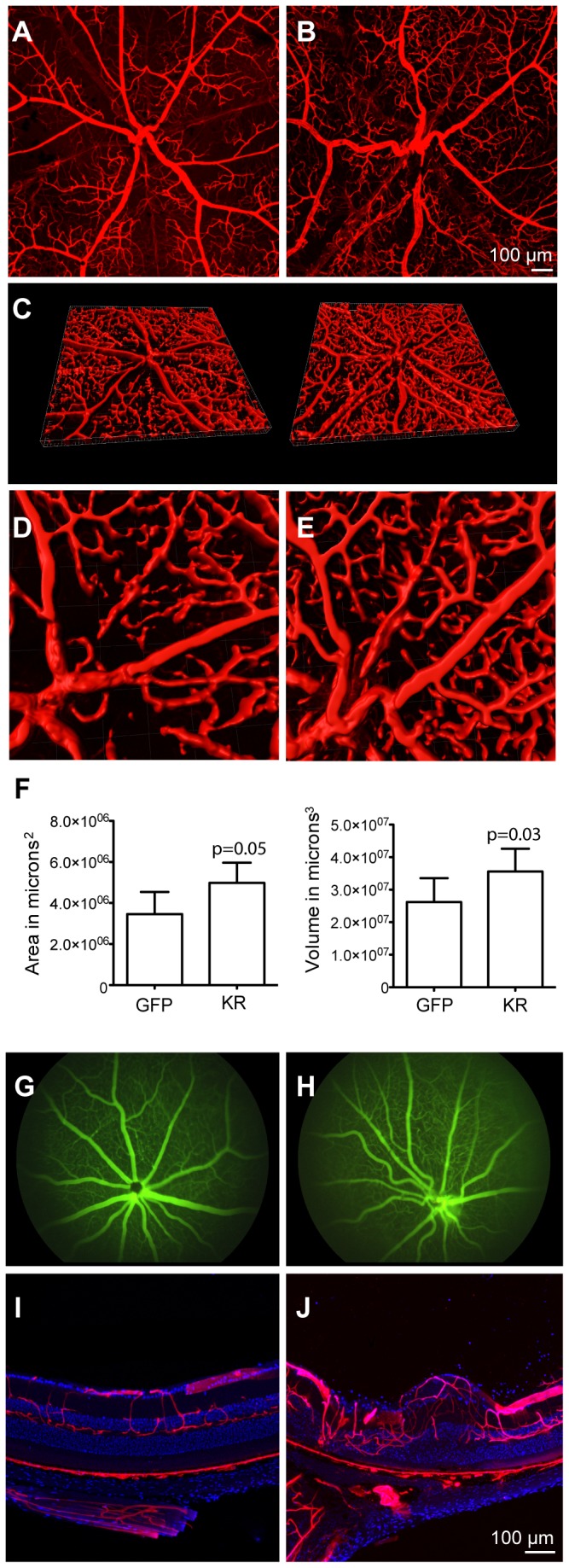
Müller glia, the primary glial cell in the retina, provide structural and metabolic support for neurons and are essential for retinal integrity. Müller cells are closely involved in many retinal degenerative diseases, including macular telangiectasia type 2, in which impairment of central vision may be linked to a primary defect in Müller glia. Here, we used an engineered, Müller-specific variant of AAV, called ShH10, to deliver a photo-inducibly toxic protein, KillerRed, to Müller cells in the mouse retina. We characterized the results of specific ablation of these cells on visual function and retinal structure. ShH10-KillerRed expression was obtained following intravitreal injection and eyes were then irradiated with green light to induce toxicity. Induction of KillerRed led to loss of Müller cells and a concomitant decrease of Müller cell markers glutamine synthetase and cellular retinaldehyde-binding protein, reduction of rhodopsin and cone opsin, and upregulation of glial fibrillary acidic protein. Loss of Müller cells also resulted in retinal disorganization, including thinning of the outer nuclear layer and the photoreceptor inner and outer segments. High resolution imaging of thin sections revealed displacement of photoreceptors from the ONL, formation of rosette-like structures and the presence of phagocytic cells. Furthermore, Müller cell ablation resulted in increased area and volume of retinal blood vessels, as well as the formation of tortuous blood vessels and vascular leakage. Electrophysiologic measures demonstrated reduced retinal function, evident in decreased photopic and scotopic electroretinogram amplitudes. These results show that loss of Müller cells can cause progressive retinal degenerative disease, and suggest that AAV delivery of an inducibly toxic protein in Müller cells may be useful to create large animal models of retinal dystrophies.
Conflict of interest statement
Figures







References
-
- Tout S, Chan-Ling T, Holländer H, Stone J (1993) The role of Müller cells in the formation of the blood-retinal barrier. Neuroscience 55: 291–301. doi:10.1016/0306-4522(93)90473-S. PubMed: 8350991. - DOI - PubMed
-
- Nishikiori N, Osanai M, Chiba H, Kojima T, Mitamura Y et al. (2007) Glial cell-derived cytokines attenuate the breakdown of vascular integrity in diabetic retinopathy. Diabetes 56: 1333–1340. doi:10.2337/db06-1431. PubMed: 17470563. - DOI - PubMed
-
- Reichenbach AA, Bringmann AA (2013) New functions of Müller cells. Glia 61: 651–678. doi:10.1002/glia.22477. PubMed: 23440929. - DOI - PubMed
-
- Bringmann A, Pannicke T, Grosche J, Francke M, Wiedemann P et al. (2006) Müller cells in the healthy and diseased retina. Prog Retin Eye Res 25: 397–424. doi:10.1016/j.preteyeres.2006.05.003. PubMed: 16839797. - DOI - PubMed
-
- Powner MB, Gillies MC, Tretiach M, Scott A, Guymer RH et al. (2010) Perifoveal Müller Cell Depletion in a Case of Macular Telangiectasia Type 2. Ophthalmology 117: 2407–2416. doi:10.1016/j.ophtha.2010.04.001. PubMed: 20678804. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
