

Condensin I associates with structural and gene regulatory regions in vertebrate chromosomes
- PMID: 24088984
- PMCID: PMC4038772
- DOI: 10.1038/ncomms3537
Condensin I associates with structural and gene regulatory regions in vertebrate chromosomes
Abstract
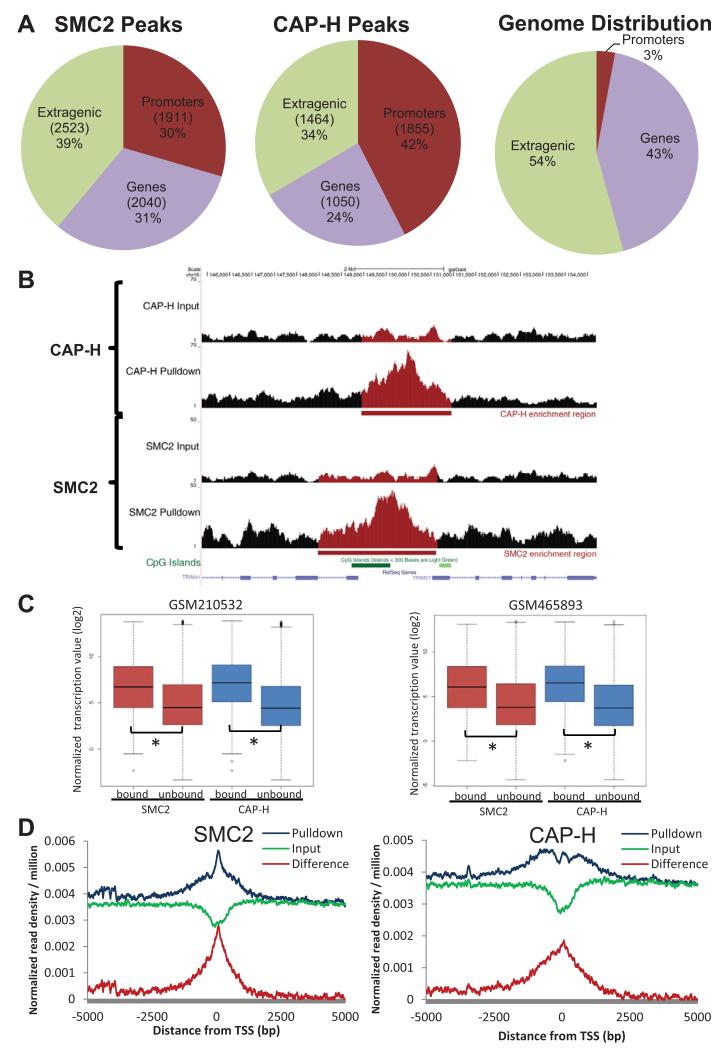
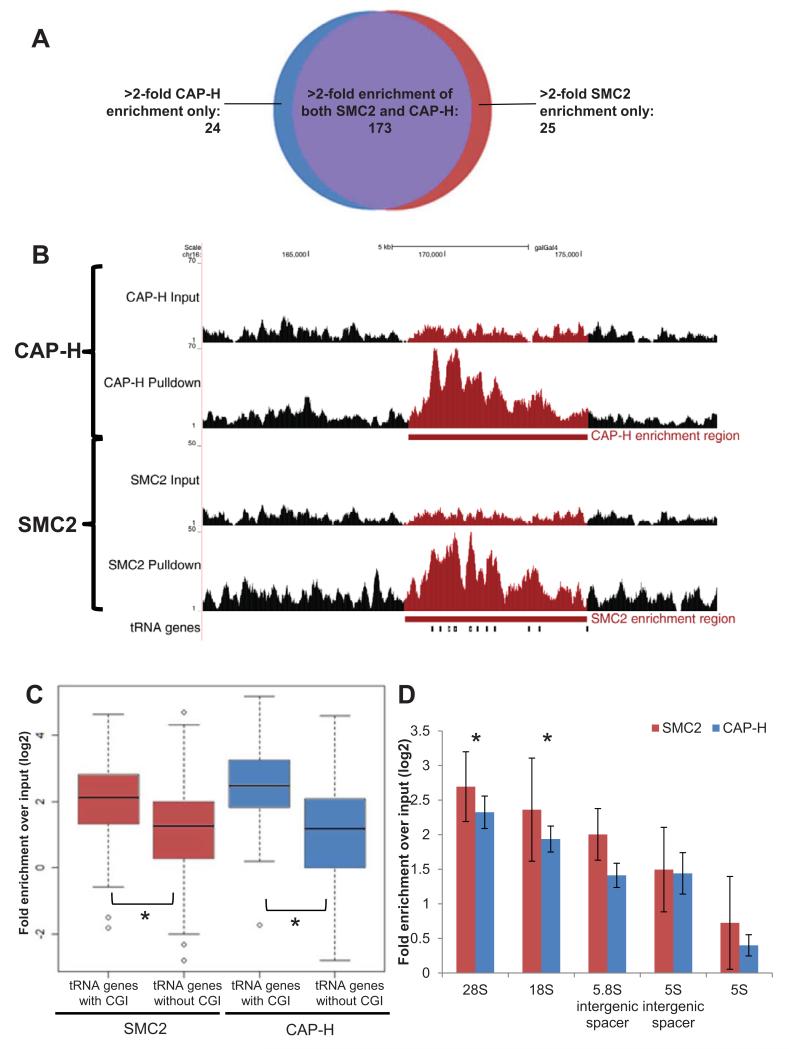
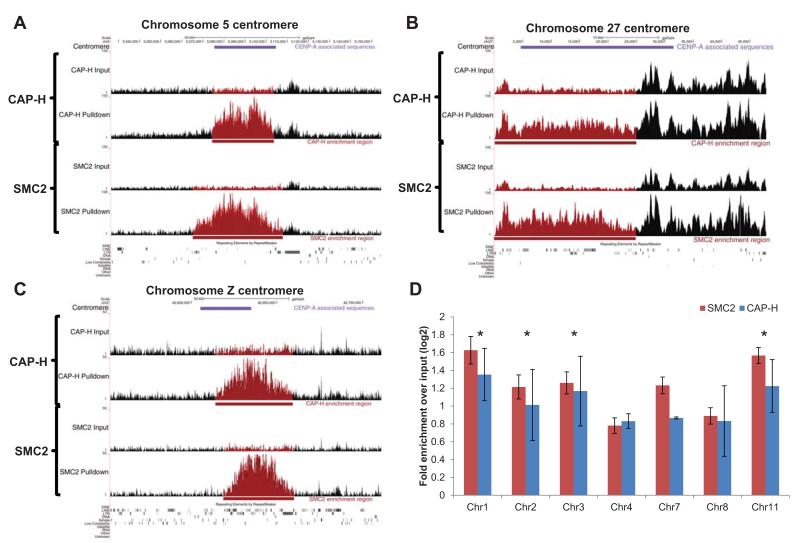
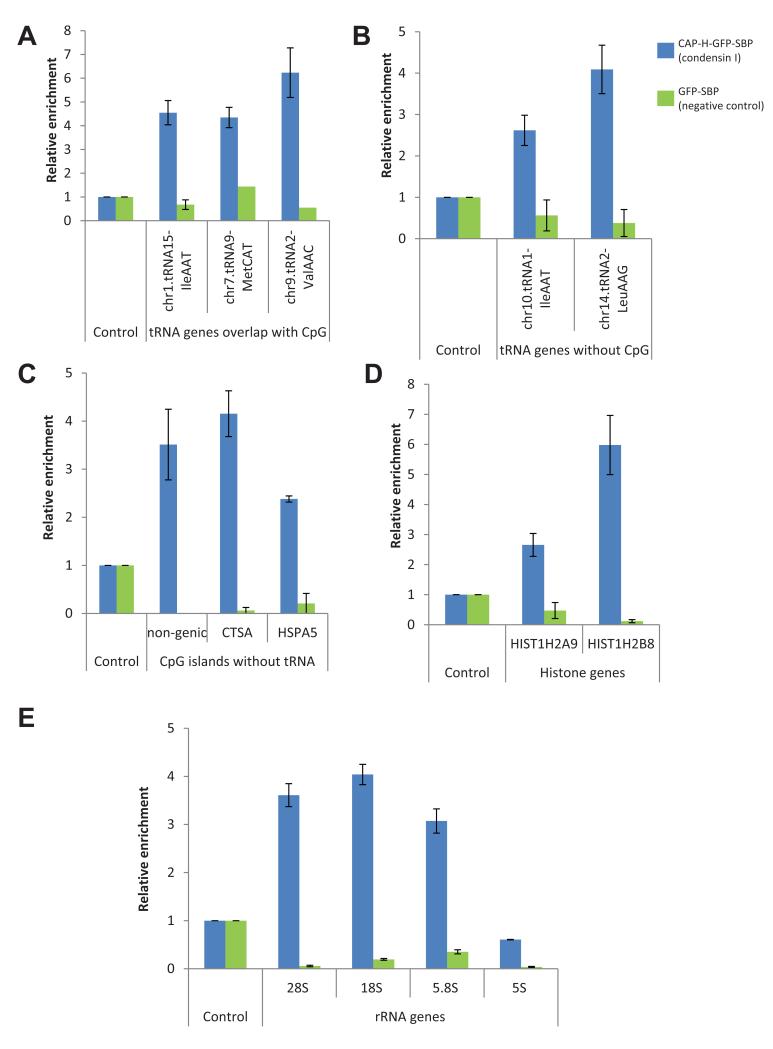
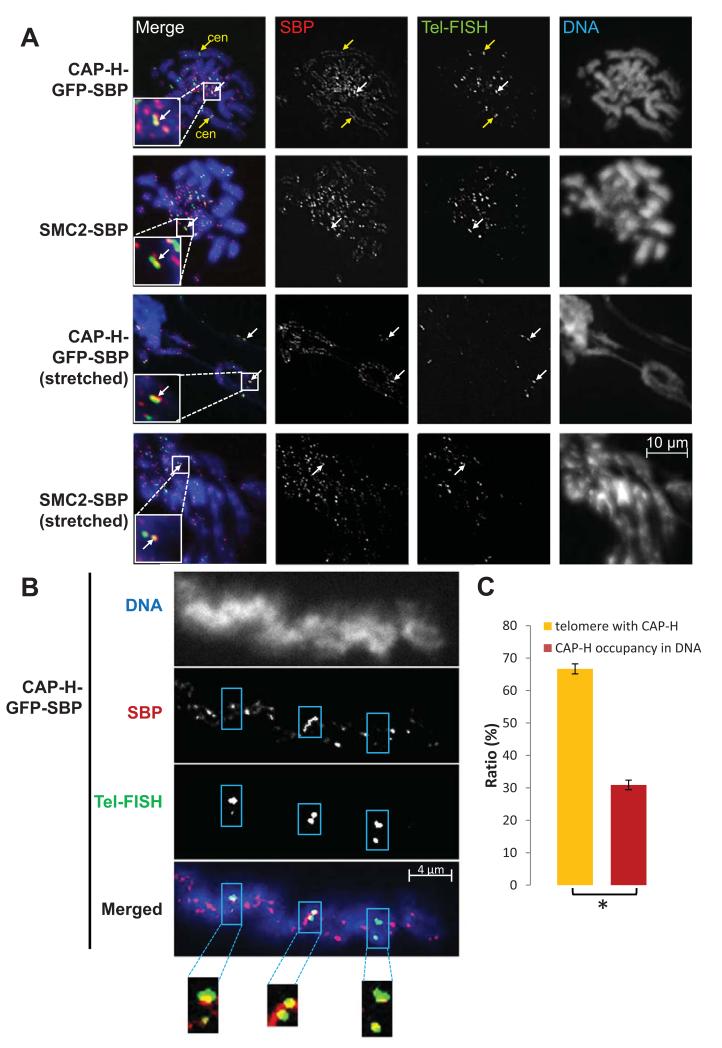
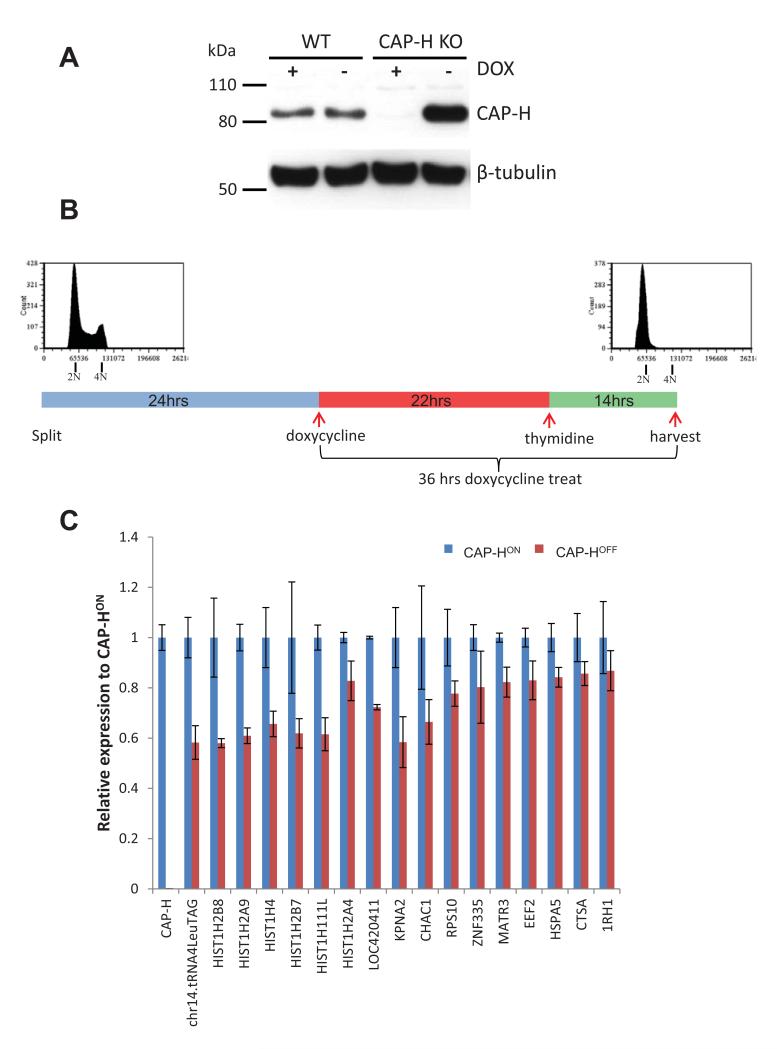
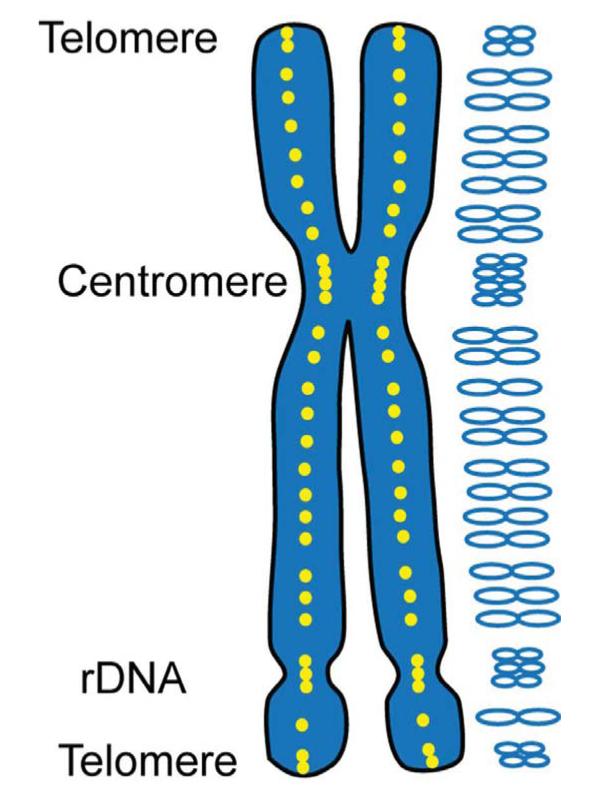
The condensin complex is essential for correct packaging and segregation of chromosomes during mitosis and meiosis in all eukaryotes. To date, the genome-wide location and the nature of condensin-binding sites have remained elusive in vertebrates. Here we report the genome-wide map of condensin I in chicken DT40 cells. Unexpectedly, we find that condensin I binds predominantly to promoter sequences in mitotic cells. We also find a striking enrichment at both centromeres and telomeres, highlighting the importance of the complex in chromosome segregation. Taken together, the results show that condensin I is largely absent from heterochromatic regions. This map of the condensin I binding sites on the chicken genome reveals that patterns of condensin distribution on chromosomes are conserved from prokaryotes, through yeasts to vertebrates. Thus in three kingdoms of life, condensin is enriched on promoters of actively transcribed genes and at loci important for chromosome segregation.
Figures
References
-
- Hirano T. At the heart of the chromosome: SMC proteins in action. Nat Rev Mol Cell Biol. 2006;7:311–322. - PubMed
-
- Saitoh N, Goldberg I, Earnshaw WC. The SMC proteins and the coming of age of the chromosome scaffold hypothesis. Bioessays. 1995;17:759–766. doi:10.1002/bies.950170905. - PubMed
-
- Aragon L, Martinez-Perez E, Merkenschlager M. Condensin, cohesin and the control of chromatin states. Curr Opin Genet Dev. 2013 doi:10.1016/j.gde.2012.11.004. - PubMed
-
- Hirano T. Condensins: Organizing and Segregating the Genome. Current Biology. 2005;15:R265–R275. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
