

Systemic disease during Streptococcus pneumoniae acute lung infection requires 12-lipoxygenase-dependent inflammation
- PMID: 24089193
- PMCID: PMC3836588
- DOI: 10.4049/jimmunol.1300522
Systemic disease during Streptococcus pneumoniae acute lung infection requires 12-lipoxygenase-dependent inflammation
Abstract
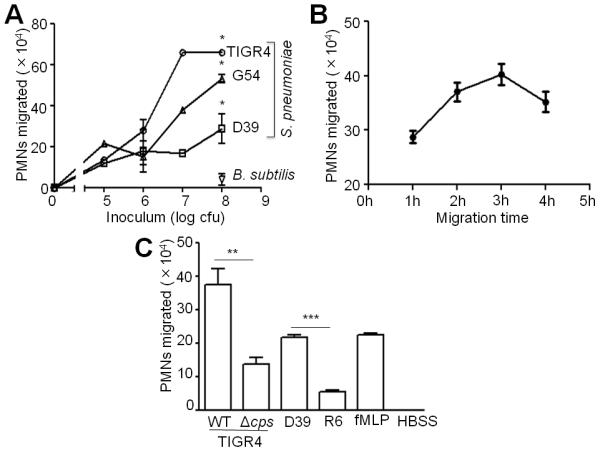
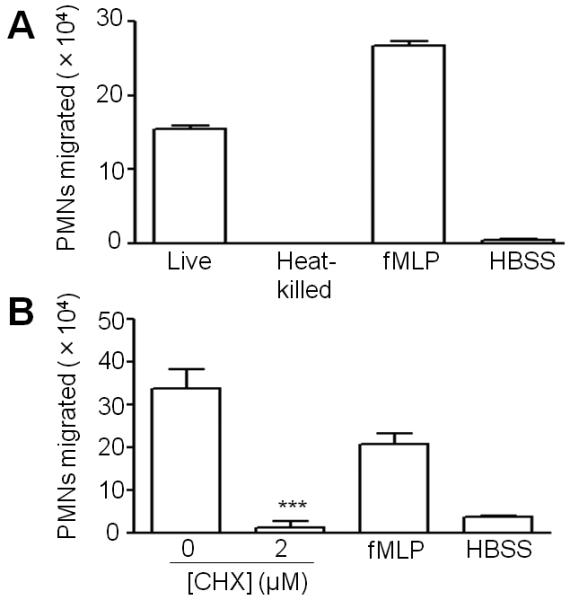
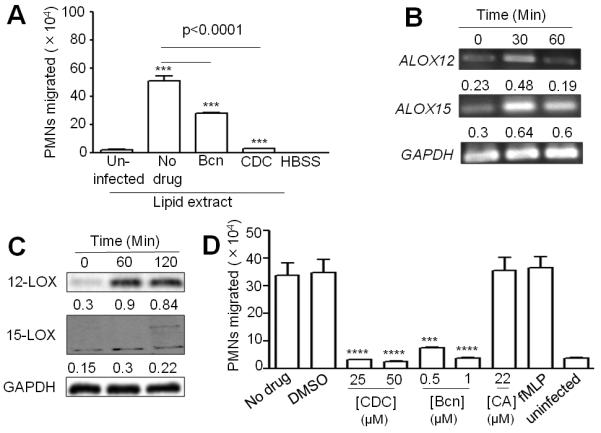
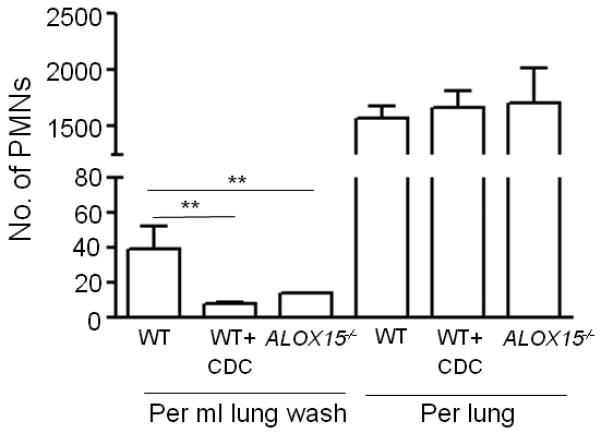
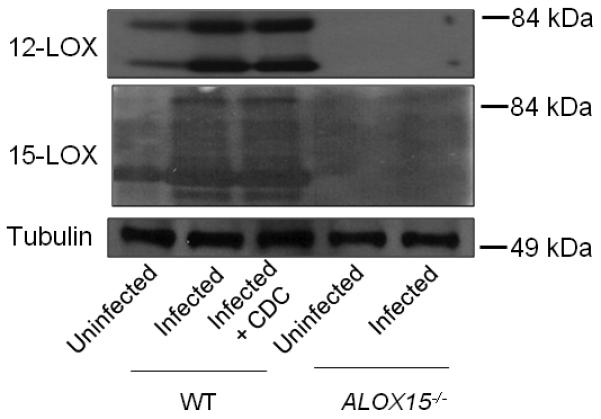
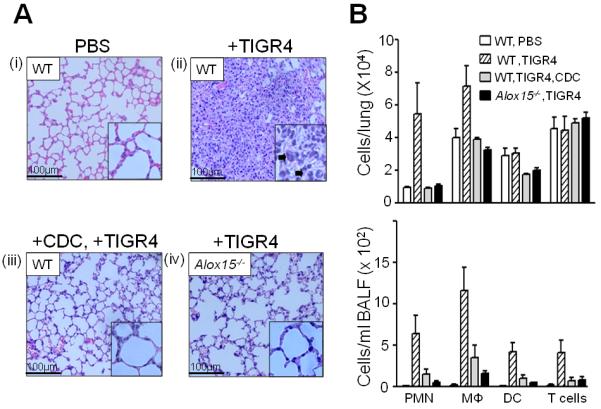
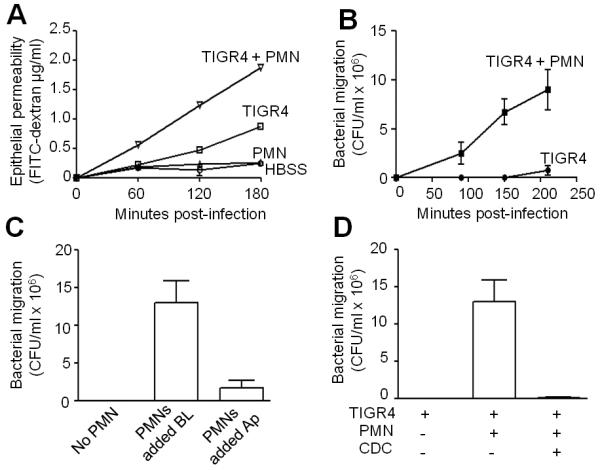
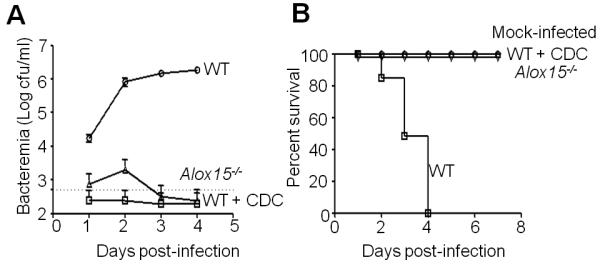
Acute pulmonary infection by Streptococcus pneumoniae is characterized by high bacterial numbers in the lung, a robust alveolar influx of polymorphonuclear cells (PMNs), and a risk of systemic spread of the bacterium. We investigated host mediators of S. pneumoniae-induced PMN migration and the role of inflammation in septicemia following pneumococcal lung infection. Hepoxilin A3 (HXA3) is a PMN chemoattractant and a metabolite of the 12-lipoxygenase (12-LOX) pathway. We observed that S. pneumoniae infection induced the production of 12-LOX in cultured pulmonary epithelium and in the lungs of infected mice. Inhibition of the 12-LOX pathway prevented pathogen-induced PMN transepithelial migration in vitro and dramatically reduced lung inflammation upon high-dose pulmonary challenge with S. pneumoniae in vivo, thus implicating HXA3 in pneumococcus-induced pulmonary inflammation. PMN basolateral-to-apical transmigration in vitro significantly increased apical-to-basolateral transepithelial migration of bacteria. Mice suppressed in the expression of 12-LOX exhibited little or no bacteremia and survived an otherwise lethal pulmonary challenge. Our data suggest that pneumococcal pulmonary inflammation is required for high-level bacteremia and systemic infection, partly by disrupting lung epithelium through 12-LOX-dependent HXA3 production and subsequent PMN transepithelial migration.
Figures
References
-
- Lynch JP, 3rd, Zhanel GG. Streptococcus pneumoniae: epidemiology and risk factors, evolution of antimicrobial resistance, and impact of vaccines. Curr. Opin. Pulm. Med. 2010;16:217–225. - PubMed
-
- Mulholland K. Strategies for the control of pneumococcal diseases. Vaccine. 1999;17(Suppl 1):S79–84. - PubMed
-
- Doerschuk CM, Markos J, Coxson HO, English D, Hogg JC. Quantitation of neutrophil migration in acute bacterial pneumonia in rabbits. J. Appl. Physiol. 1994;77:2593–2599. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
