

Low selection pressure aids the evolution of cooperative ribozyme mutations in cells
- PMID: 24089519
- PMCID: PMC3829158
- DOI: 10.1074/jbc.M113.511469
Low selection pressure aids the evolution of cooperative ribozyme mutations in cells
Abstract
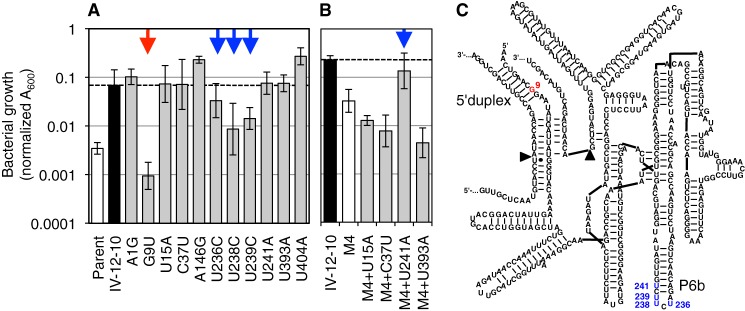
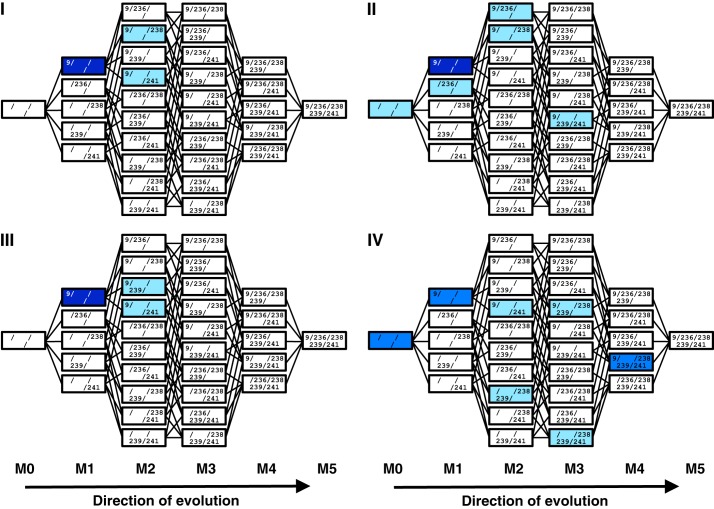
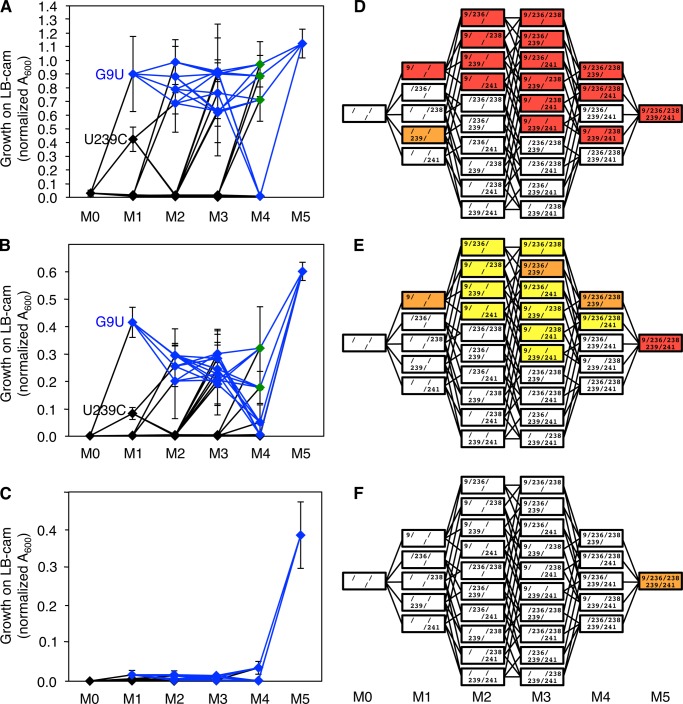
Understanding the evolution of functional RNA molecules is important for our molecular understanding of biology. Here we tested experimentally how two evolutionary parameters, selection pressure and recombination, influenced the evolution of an evolving RNA population. This was done using four parallel evolution experiments that employed low or gradually increasing selection pressure, and recombination events either at the end or dispersed throughout the evolution. As model system, a trans-splicing group I intron ribozyme was evolved in Escherichia coli cells over 12 rounds of selection and amplification, including mutagenesis and recombination. The low selection pressure resulted in higher efficiency of the evolved ribozyme populations, whereas differences in recombination did not have a strong effect. Five mutations were responsible for the highest efficiency. The first mutation swept quickly through all four evolving populations, whereas the remaining four mutations accumulated later and more efficiently under low selection pressure. To determine why low selection pressure aided this evolution, all evolutionary intermediates between the wild type and the 5-mutation variant were constructed, and their activities at three different selection pressures were determined. The resulting fitness profiles showed a high cooperativity among the four late mutations, which can explain why high selection pressure led to inefficient evolution. These results show experimentally how low selection pressure can benefit the evolution of cooperative mutations in functional RNAs.
Keywords: Cooperativity; Escherichia coli; Evolution; RNA Catalysis; Ribozyme.
Figures
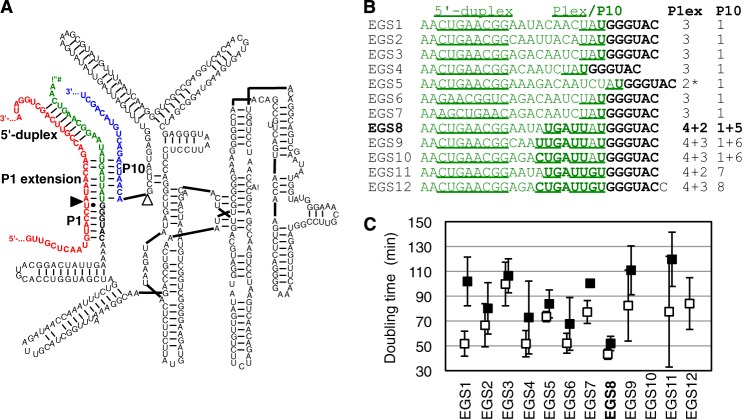
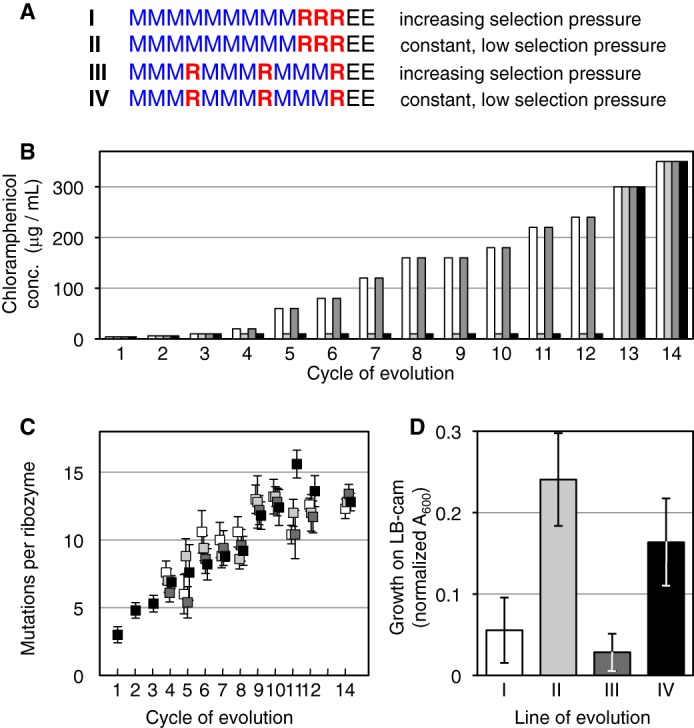
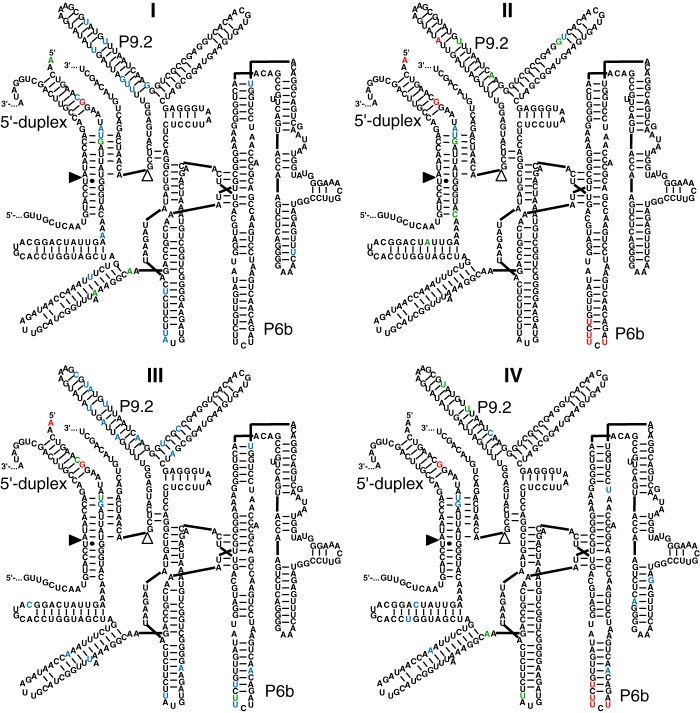
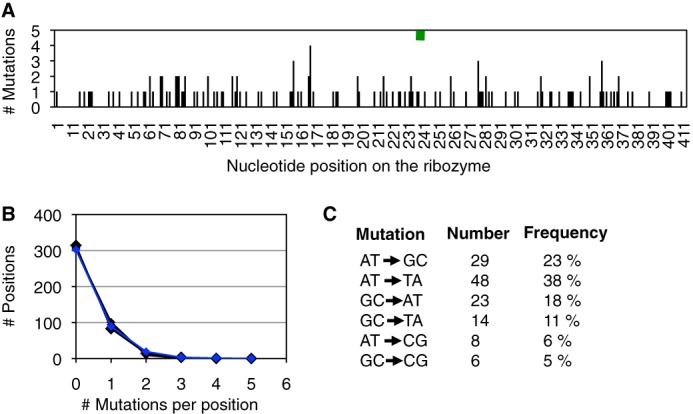
References
-
- Noller H. F., Hoffarth V., Zimniak L. (1992) Unusual resistance of peptidyl transferase to protein extraction procedures. Science 256, 1416–1419 - PubMed
-
- Nissen P., Hansen J., Ban N., Moore P. B., Steitz T. A. (2000) The structural basis of ribosome activity in peptide bond synthesis. Science 289, 920–930 - PubMed
-
- Guerrier-Takada C., Gardiner K., Marsh T., Pace N., Altman S. (1983) The RNA moiety of ribonuclease P is the catalytic subunit of the enzyme. Cell 35, 849–857 - PubMed
-
- Padgett R. A., Grabowski P. J., Konarska M. M., Seiler S., Sharp P. A. (1986) Splicing of messenger RNA precursors. Annu. Rev. Biochem. 55, 1119–1150 - PubMed
-
- Valadkhan S. (2005) snRNAs as the catalysts of pre-mRNA splicing. Curr. Opin. Chem. Biol. 9, 603–608 - PubMed
