

Human eukaryotic initiation factor 4G (eIF4G) protein binds to eIF3c, -d, and -e to promote mRNA recruitment to the ribosome
- PMID: 24092755
- PMCID: PMC3829144
- DOI: 10.1074/jbc.M113.517011
Human eukaryotic initiation factor 4G (eIF4G) protein binds to eIF3c, -d, and -e to promote mRNA recruitment to the ribosome
Abstract
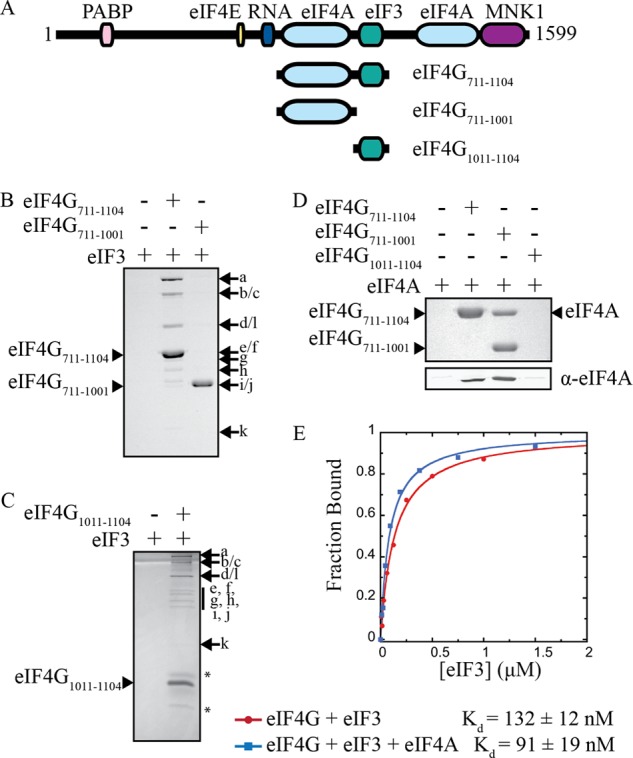
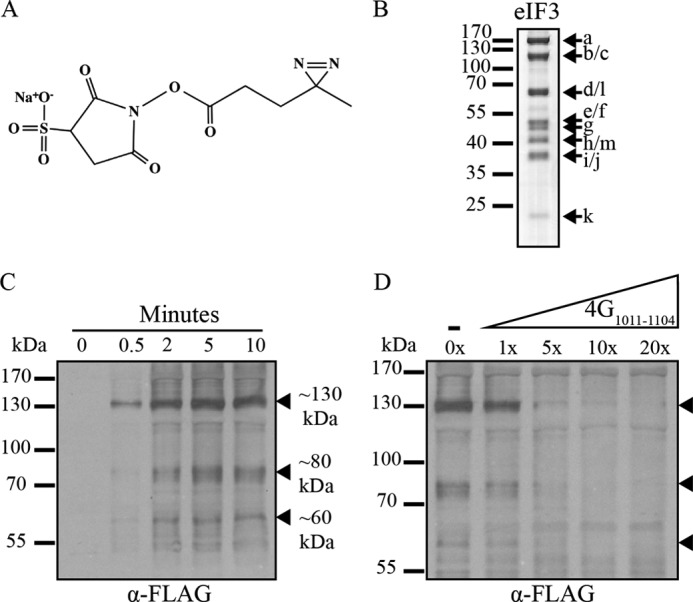
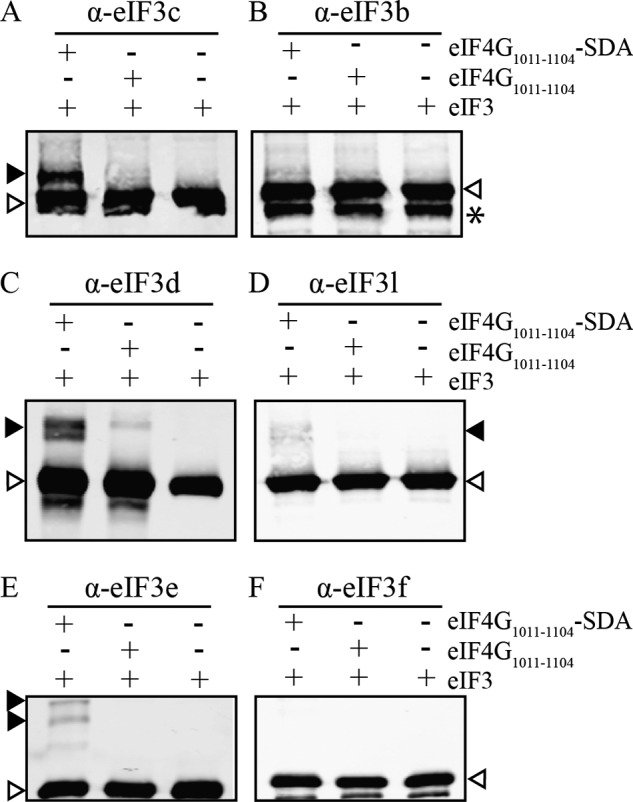
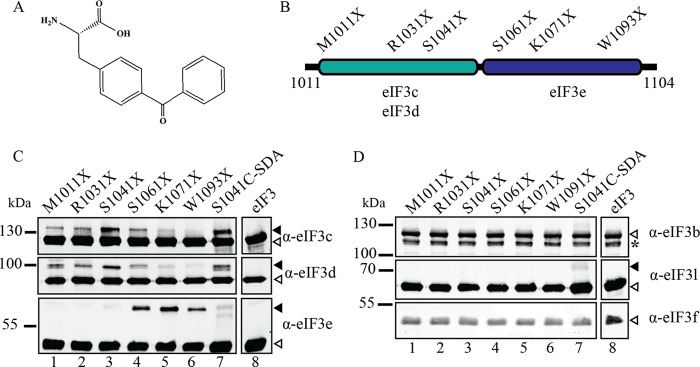
Recruitment of mRNA to the 40S ribosomal subunit requires the coordinated interaction of a large number of translation initiation factors. In mammals, the direct interaction between eukaryotic initiation factor 4G (eIF4G) and eIF3 is thought to act as the molecular bridge between the mRNA cap-binding complex and the 40S subunit. A discrete ∼90 amino acid domain in eIF4G is responsible for binding to eIF3, but the identity of the eIF3 subunit(s) involved is less clear. The eIF3e subunit has been shown to directly bind eIF4G, but the potential role of other eIF3 subunits in stabilizing this interaction has not been investigated. It is also not clear if the eIF4A helicase plays a role in stabilizing the interaction between eIF4G and eIF3. Here, we have used a fluorescence anisotropy assay to demonstrate that eIF4G binds to eIF3 independently of eIF4A binding to the middle region of eIF4G. By using a site-specific cross-linking approach, we unexpectedly show that the eIF4G-binding surface in eIF3 is comprised of the -c, -d and -e subunits. Screening multiple cross-linker positions reveals that eIF4G contains two distinct eIF3-binding subdomains within the previously identified eIF3-binding domain. Finally, by employing an eIF4G-dependent translation assay, we establish that both of these subdomains are required for efficient mRNA recruitment to the ribosome and stimulate translation. Our study reveals unexpected complexity to the eIF3-eIF4G interaction that provides new insight into the regulation of mRNA recruitment to the human ribosome.
Keywords: Initiation; Protein Cross-linking; Protein-Protein Interactions; Ribosomes; Translation; eIF3; eIF4G; mRNA.
Figures
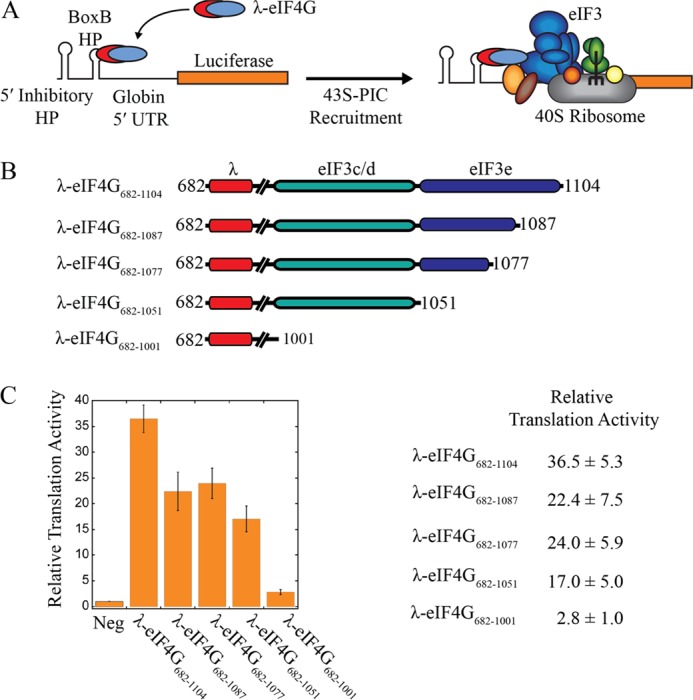
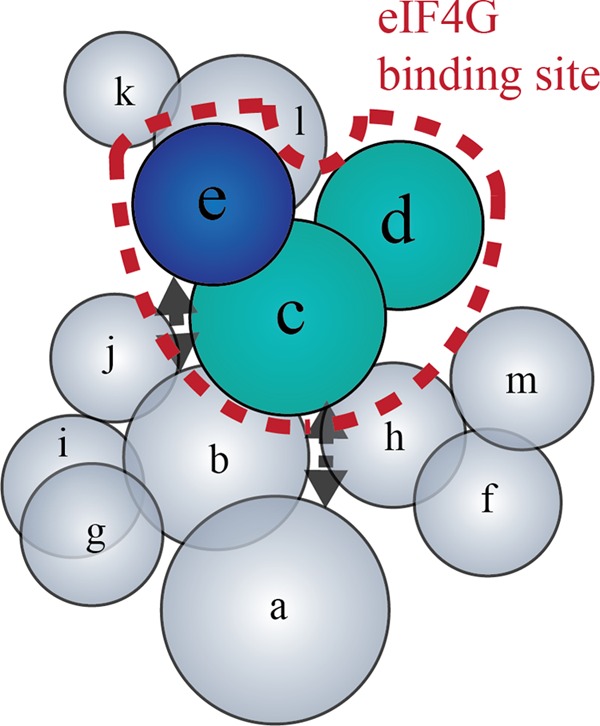
References
-
- Fraser C. S. (2009) The molecular basis of translational control. Prog. Mol. Biol. Transl. Sci. 90, 1–51 - PubMed
-
- Gingras A. C., Raught B., Sonenberg N. (1999) eIF4 initiation factors: effectors of mRNA recruitment to ribosomes and regulators of translation. Annu. Rev. Biochem. 68, 913–963 - PubMed
-
- Parsyan A., Svitkin Y., Shahbazian D., Gkogkas C., Lasko P., Merrick W. C., Sonenberg N. (2011) mRNA helicases: the tacticians of translational control. Nat. Rev. Mol. Cell Biol. 12, 235–245 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
