

Bronchus-associated lymphoid tissue in pulmonary hypertension produces pathologic autoantibodies
- PMID: 24093638
- PMCID: PMC3863738
- DOI: 10.1164/rccm.201302-0403OC
Bronchus-associated lymphoid tissue in pulmonary hypertension produces pathologic autoantibodies
Abstract
Rationale: Autoimmunity has long been associated with pulmonary hypertension. Bronchus-associated lymphoid tissue plays important roles in antigen sampling and self-tolerance during infection and inflammation.
Objectives: We reasoned that activated bronchus-associated lymphoid tissue would be evident in rats with pulmonary hypertension, and that loss of self-tolerance would result in production of pathologic autoantibodies that drive vascular remodeling.
Methods: We used animal models, histology, and gene expression assays to evaluate the role of bronchus-associated lymphoid tissue in pulmonary hypertension.
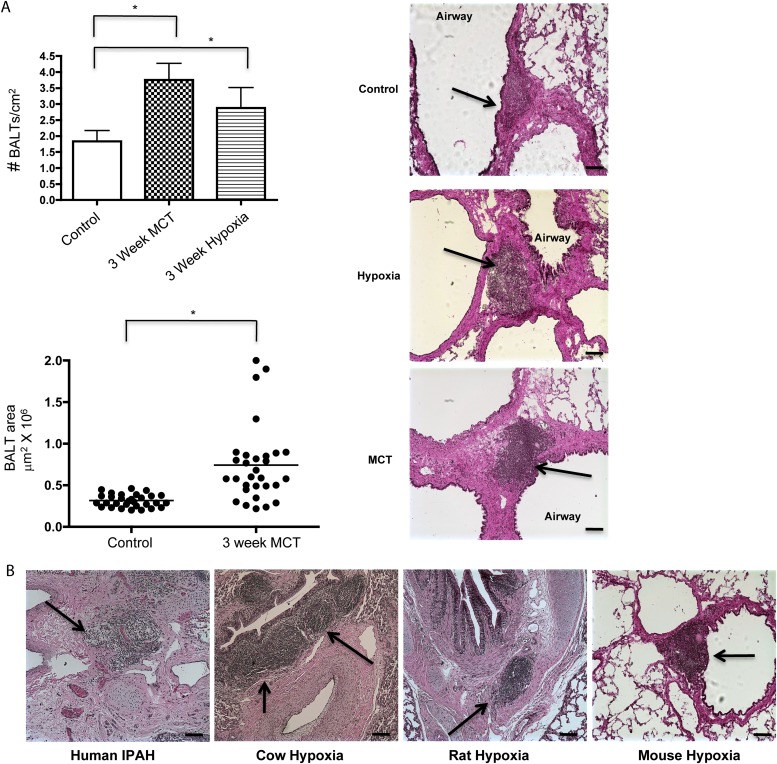
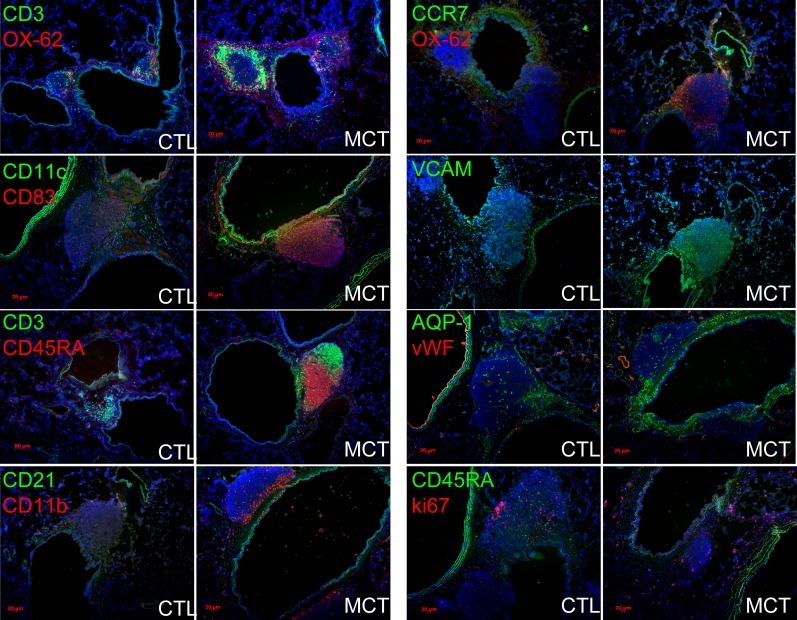
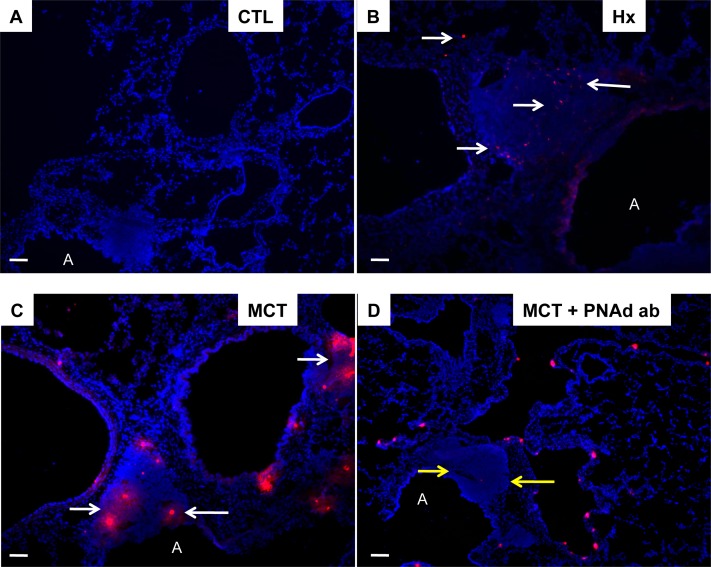
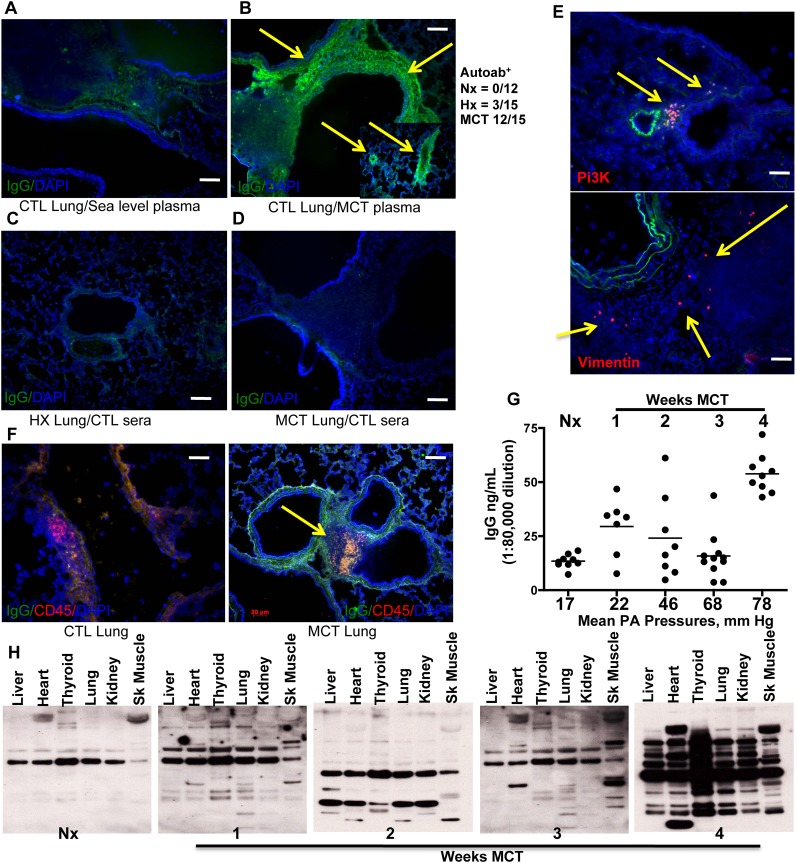
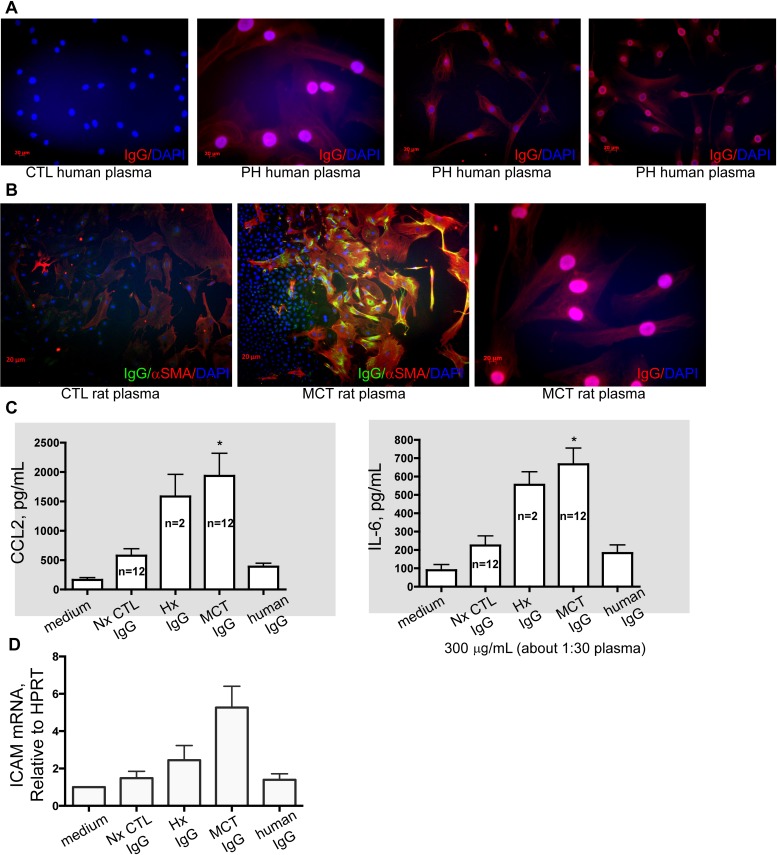
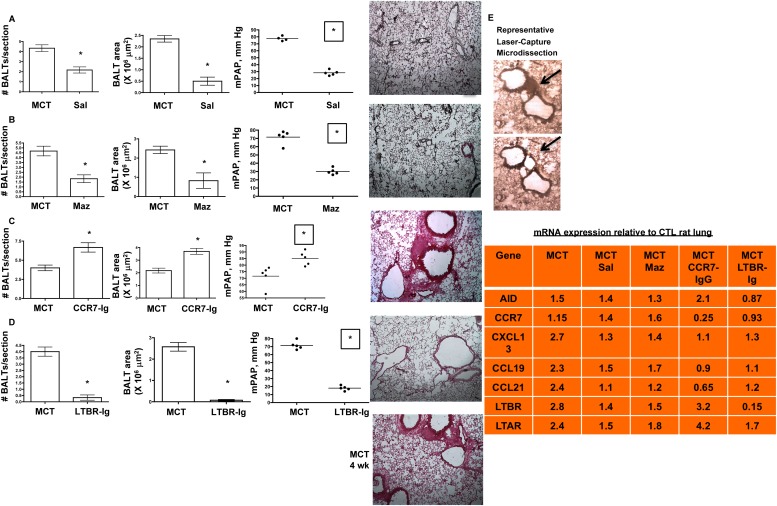
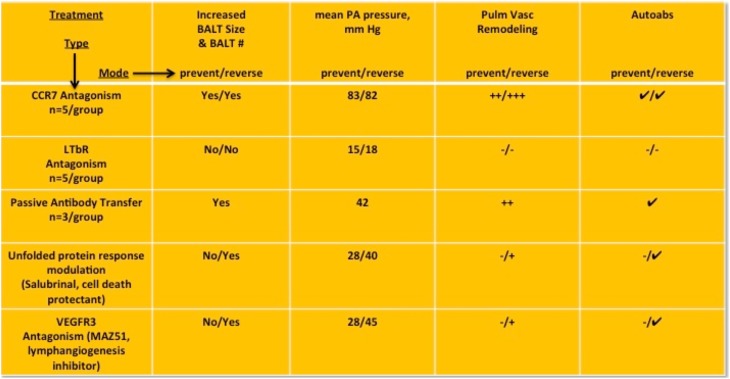
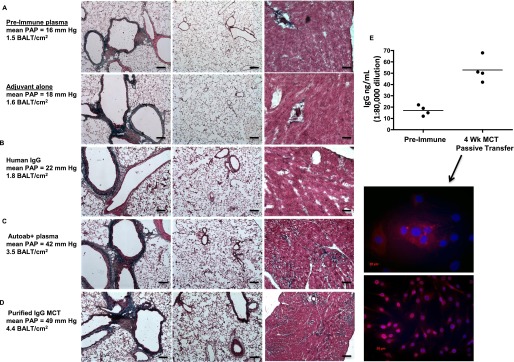
Measurements and main results: Bronchus-associated lymphoid tissue was more numerous, larger, and more active in pulmonary hypertension compared with control animals. We found dendritic cells in and around lymphoid tissue, which were composed of CD3(+) T cells over a core of CD45RA(+) B cells. Antirat IgG and plasma from rats with pulmonary hypertension decorated B cells in lymphoid tissue, resistance vessels, and adventitia of large vessels. Lymphoid tissue in diseased rats was vascularized by aquaporin-1(+) high endothelial venules and vascular cell adhesion molecule-positive vessels. Autoantibodies are produced in bronchus-associated lymphoid tissue and, when bound to pulmonary adventitial fibroblasts, change their phenotype to one that may promote inflammation. Passive transfer of autoantibodies into rats caused pulmonary vascular remodeling and pulmonary hypertension. Diminution of lymphoid tissue reversed pulmonary hypertension, whereas immunologic blockade of CCR7 worsened pulmonary hypertension and hastened its onset.
Conclusions: Bronchus-associated lymphoid tissue expands in pulmonary hypertension and is autoimmunologically active. Loss of self-tolerance contributes to pulmonary vascular remodeling and pulmonary hypertension. Lymphoid tissue-directed therapies may be beneficial in treating pulmonary hypertension.
Figures
References
-
- Simonneau G, Robbins IM, Beghetti M, Channick RN, Delcroix M, Denton CP, Elliott CG, Gaine SP, Gladwin MT, Jing ZC, et al. Updated clinical classification of pulmonary hypertension. J Am Coll Cardiol. 2009;54:S43–S54. - PubMed
-
- Tuder RM, Abman SH, Braun T, Capron F, Stevens T, Thistlethwaite PA, Haworth SG. Development and pathology of pulmonary hypertension. J Am Coll Cardiol. 2009;54:S3–S9. - PubMed
-
- Hassoun PM, Mouthon L, Barberà JA, Eddahibi S, Flores SC, Grimminger F, Jones PL, Maitland ML, Michelakis ED, Morrell NW, et al. Inflammation, growth factors, and pulmonary vascular remodeling. J Am Coll Cardiol. 2009;54:S10–19. - PubMed
-
- Foo SY, Phipps S. Regulation of inducible BALT formation and contribution to immunity and pathology. Mucosal Immunol. 2010;3:537–544. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
