

The role of vimentin intermediate filaments in cortical and cytoplasmic mechanics
- PMID: 24094397
- PMCID: PMC3791300
- DOI: 10.1016/j.bpj.2013.08.037
The role of vimentin intermediate filaments in cortical and cytoplasmic mechanics
Abstract
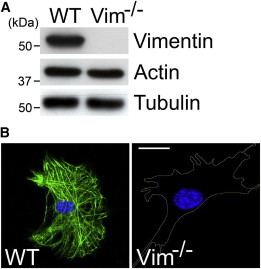
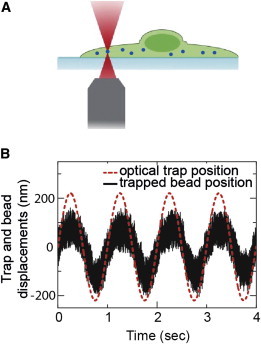
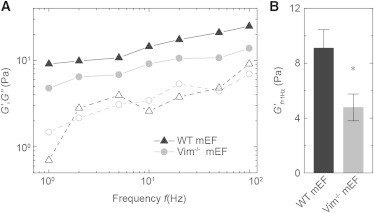
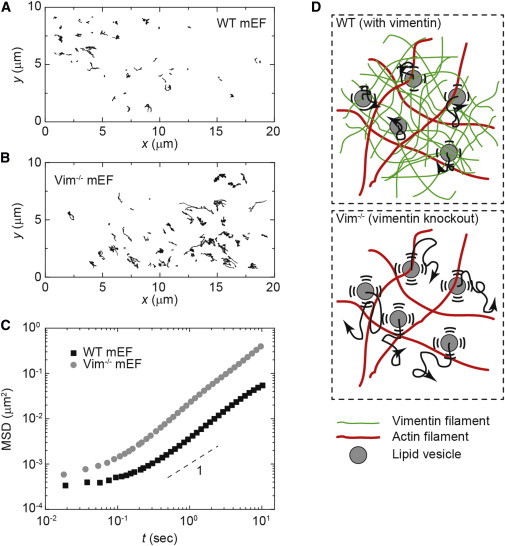
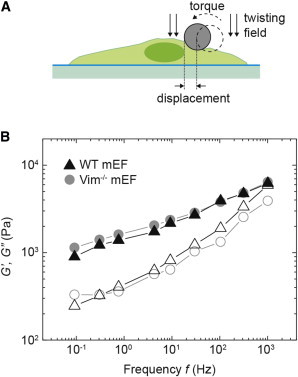
The mechanical properties of a cell determine many aspects of its behavior, and these mechanics are largely determined by the cytoskeleton. Although the contribution of actin filaments and microtubules to the mechanics of cells has been investigated in great detail, relatively little is known about the contribution of the third major cytoskeletal component, intermediate filaments (IFs). To determine the role of vimentin IF (VIF) in modulating intracellular and cortical mechanics, we carried out studies using mouse embryonic fibroblasts (mEFs) derived from wild-type or vimentin(-/-) mice. The VIFs contribute little to cortical stiffness but are critical for regulating intracellular mechanics. Active microrheology measurements using optical tweezers in living cells reveal that the presence of VIFs doubles the value of the cytoplasmic shear modulus to ∼10 Pa. The higher levels of cytoplasmic stiffness appear to stabilize organelles in the cell, as measured by tracking endogenous vesicle movement. These studies show that VIFs both increase the mechanical integrity of cells and localize intracellular components.
Copyright © 2013 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures
Comment in
-
Mechanical players-The role of intermediate filaments in cell mechanics and organization.Biophys J. 2013 Oct 15;105(8):1733-4. doi: 10.1016/j.bpj.2013.08.050. Biophys J. 2013. PMID: 24138847 Free PMC article. No abstract available.
References
-
- Alberts B., Johnson A., Lewis J., Raff M., Roberts K., Walter P. Garland Science; New York: 2007. Molecular Biology of the Cell.
-
- Carlier M.F. Role of nucleotide hydrolysis in the dynamics of actin filaments and microtubules. Int. Rev. Cytol. 1989;115:139–170. - PubMed
-
- Herrmann H., Bär H., Aebi U. Intermediate filaments: from cell architecture to nanomechanics. Nat. Rev. Mol. Cell Biol. 2007;8:562–573. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
