

p53-dependent gene repression through p21 is mediated by recruitment of E2F4 repression complexes
- PMID: 24096481
- PMCID: PMC4067464
- DOI: 10.1038/onc.2013.378
p53-dependent gene repression through p21 is mediated by recruitment of E2F4 repression complexes
Abstract
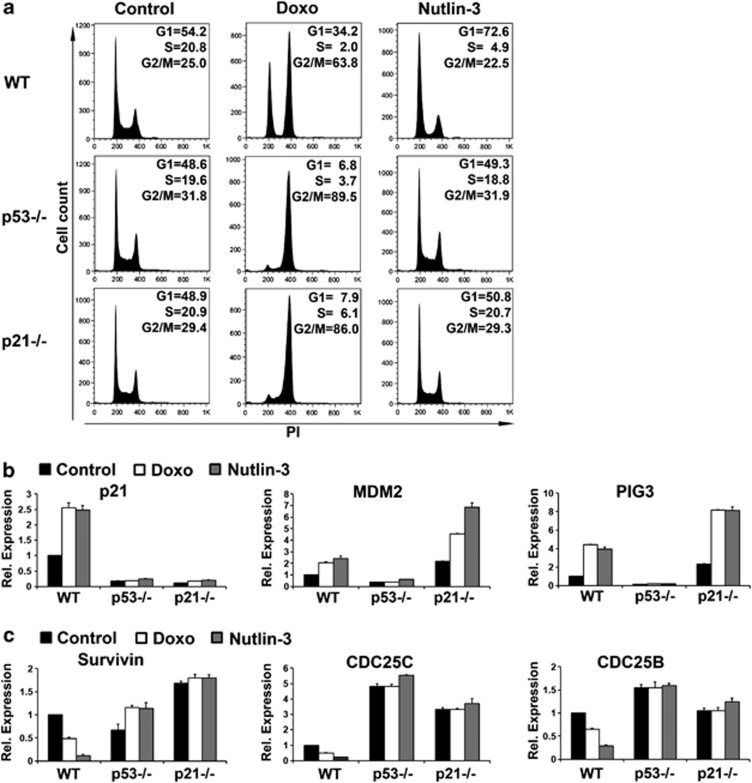
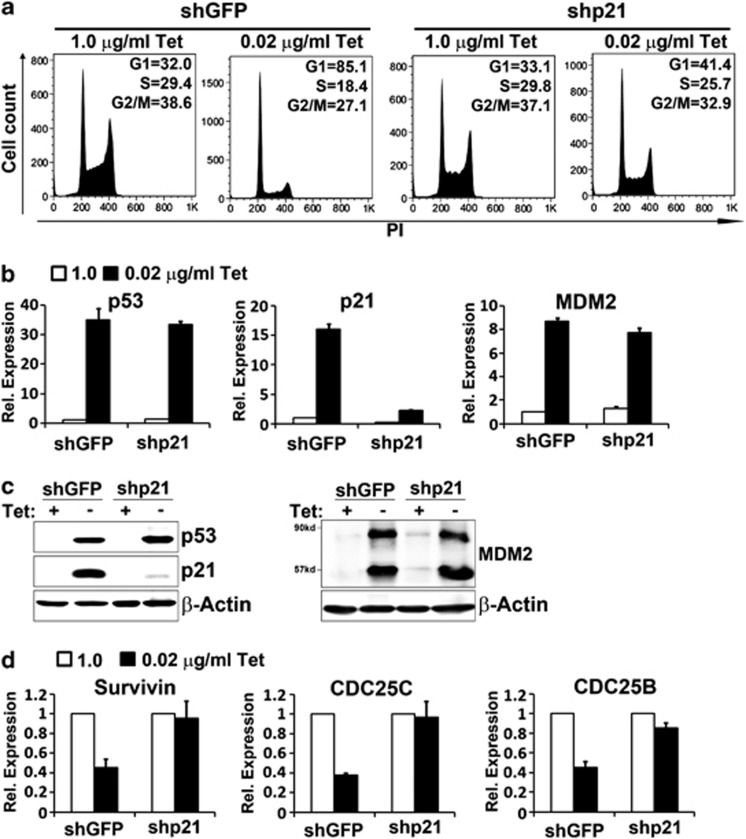
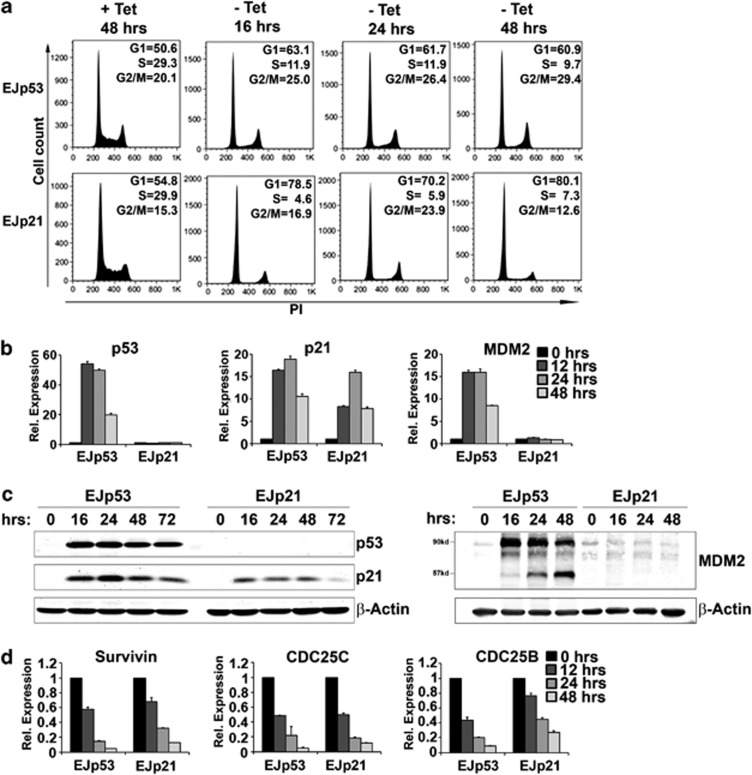
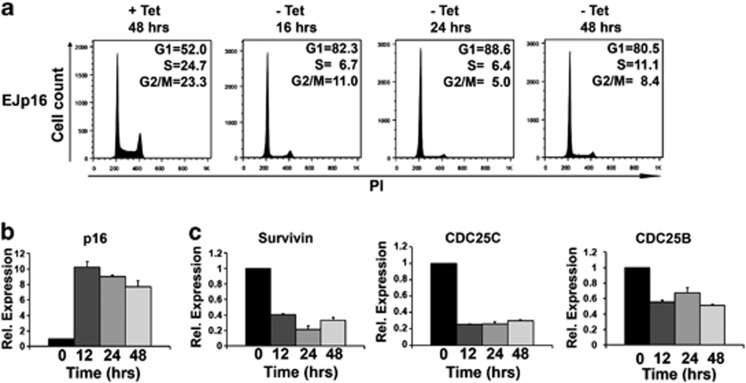
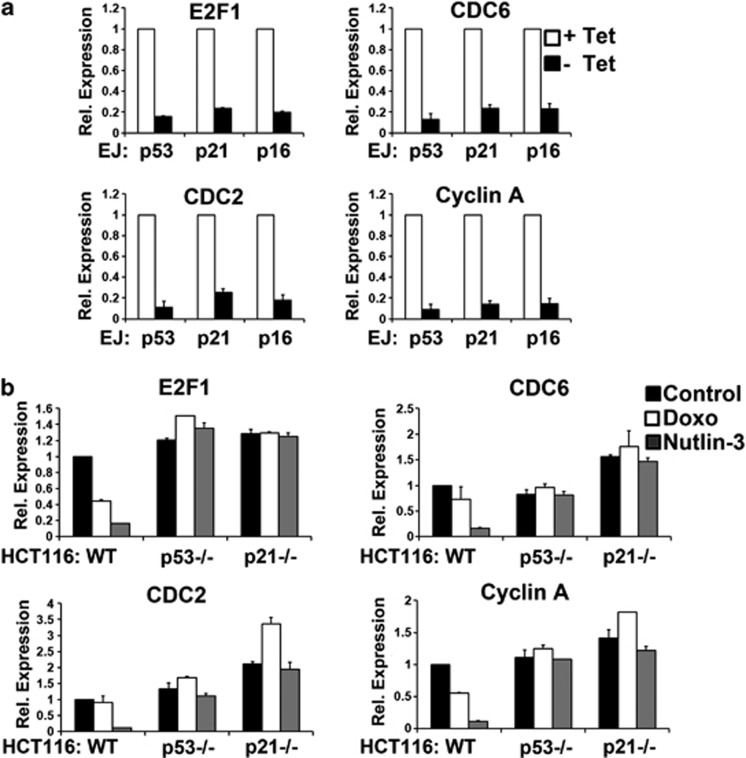
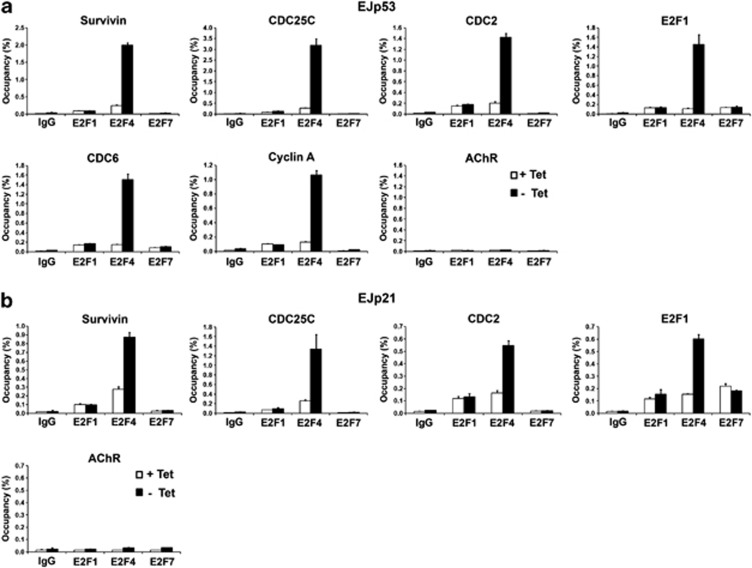
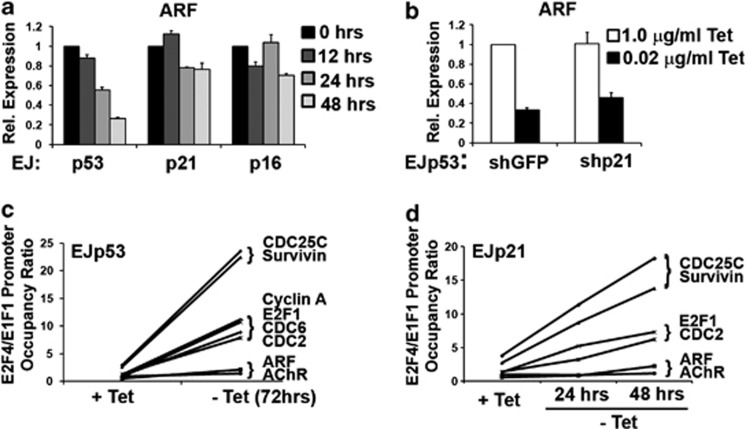
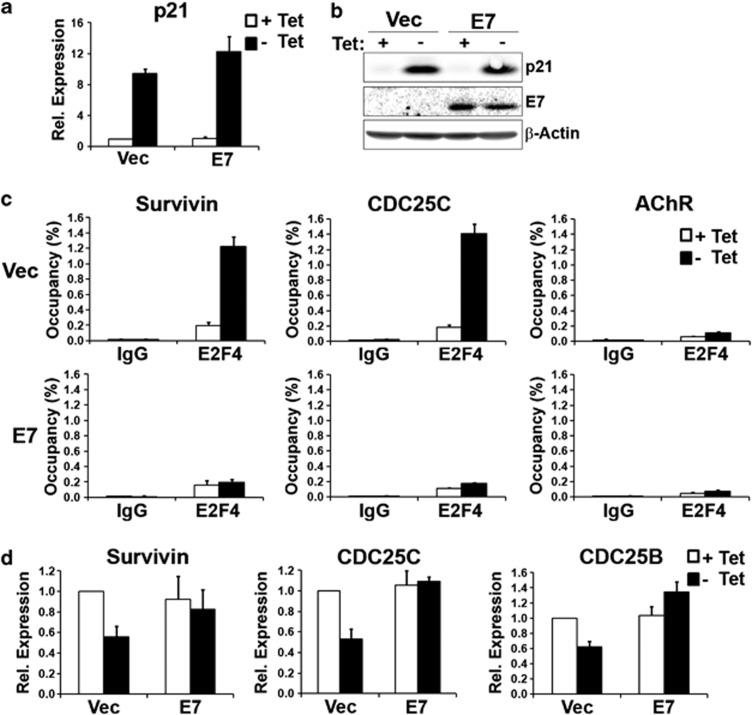
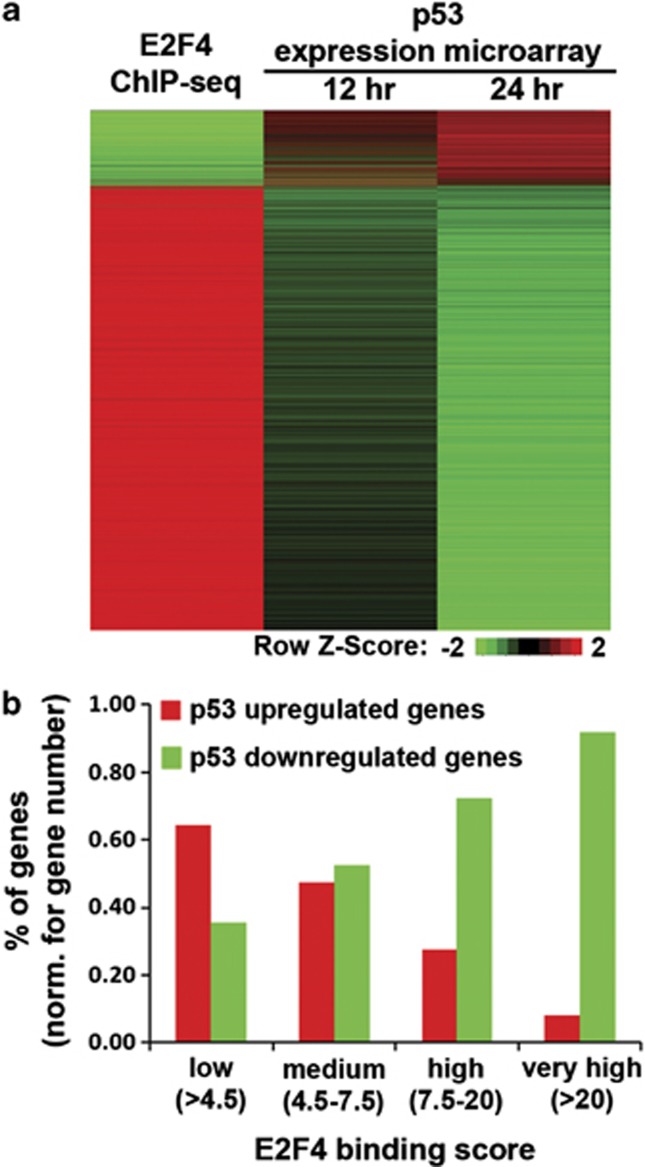
The p53 tumor suppressor protein is a major sensor of cellular stresses, and upon stabilization, activates or represses many genes that control cell fate decisions. While the mechanism of p53-mediated transactivation is well established, several mechanisms have been proposed for p53-mediated repression. Here, we demonstrate that the cyclin-dependent kinase inhibitor p21 is both necessary and sufficient for the downregulation of known p53-repression targets, including survivin, CDC25C, and CDC25B in response to p53 induction. These same targets are similarly repressed in response to p16 overexpression, implicating the involvement of the shared downstream retinoblastoma (RB)-E2F pathway. We further show that in response to either p53 or p21 induction, E2F4 complexes are specifically recruited onto the promoters of these p53-repression targets. Moreover, abrogation of E2F4 recruitment via the inactivation of RB pocket proteins, but not by RB loss of function alone, prevents the repression of these genes. Finally, our results indicate that E2F4 promoter occupancy is globally associated with p53-repression targets, but not with p53 activation targets, implicating E2F4 complexes as effectors of p21-dependent p53-mediated repression.
Figures
References
-
- Vousden KH, Prives C. Blinded by the light: the growing complexity of p53. Cell. 2009;137:413–431. - PubMed
-
- El-Deiry WS. The role of p53 in chemosensitivity and radiosensitivity. Oncogene. 2003;22:7486–7495. - PubMed
-
- Alarcon-Vargas D, Ronai Z. p53-Mdm2–the affair that never ends. Carcinogenesis. 2002;23:541–547. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
