

Imaging energy status in live cells with a fluorescent biosensor of the intracellular ATP-to-ADP ratio
- PMID: 24096541
- PMCID: PMC3852917
- DOI: 10.1038/ncomms3550
Imaging energy status in live cells with a fluorescent biosensor of the intracellular ATP-to-ADP ratio
Abstract
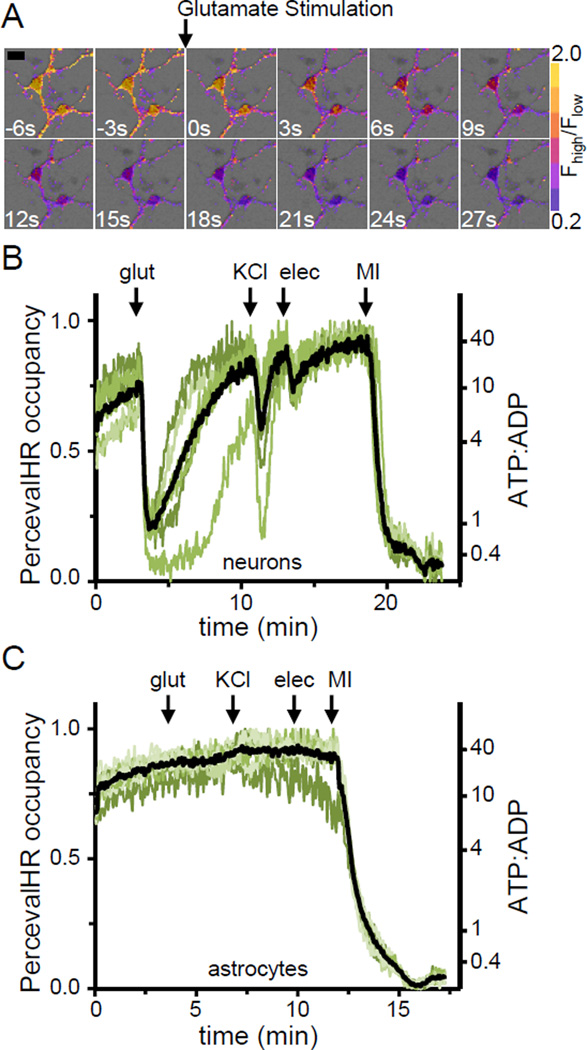
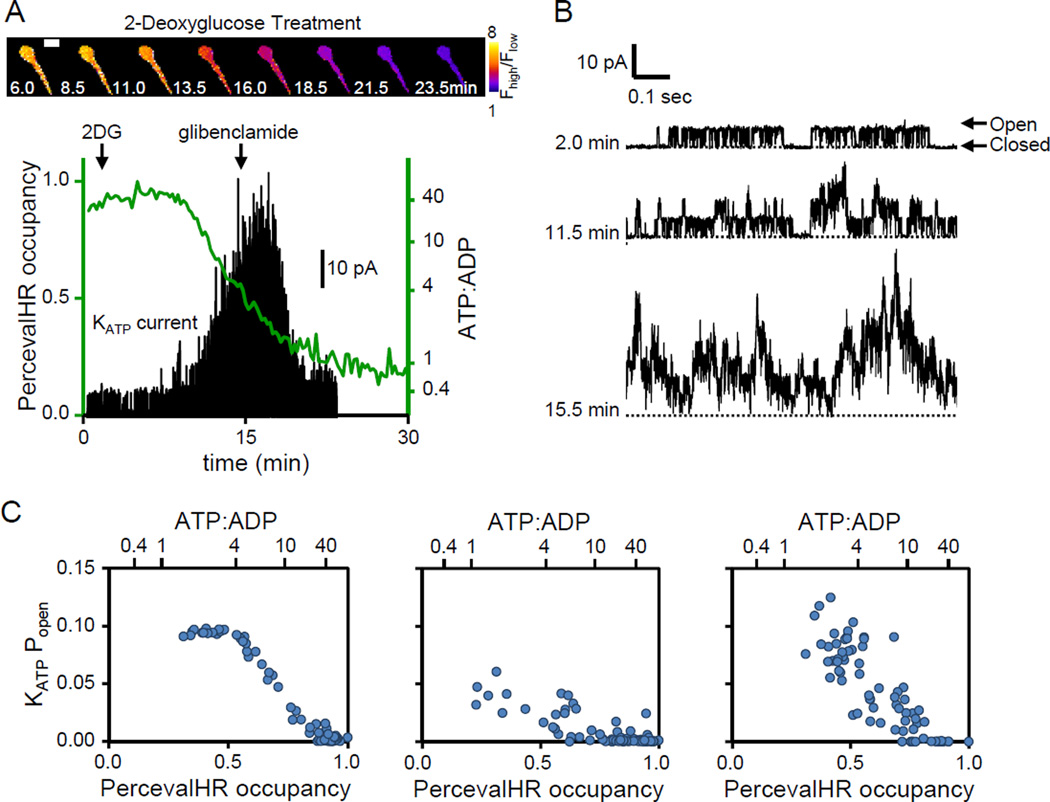
The ATP:ADP ratio is a critical parameter of cellular energy status that regulates many metabolic activities. Here we report an optimized genetically encoded fluorescent biosensor, PercevalHR, that senses the ATP:ADP ratio. PercevalHR is tuned to the range of intracellular ATP:ADP expected in mammalian cells, and it can be used with one- or two-photon microscopy in live samples. We use PercevalHR to visualize activity-dependent changes in ATP:ADP when neurons are exposed to multiple stimuli, demonstrating that it is a sensitive reporter of physiological changes in energy consumption and production. We also use PercevalHR to visualize intracellular ATP:ADP while simultaneously recording currents from ATP-sensitive potassium (KATP) channels in single cells, showing that PercevalHR enables the study of coordinated variation in ATP:ADP and KATP channel open probability in intact cells. With its ability to monitor changes in cellular energetics within seconds, PercevalHR should be a versatile tool for metabolic research.
Figures





References
-
- Veech RL, Lawson JW, Cornell NW, Krebs HA. Cytosolic phosphorylation potential. J. Biol. Chem. 1979;254:6538–6547. - PubMed
-
- Gabriel JL, Zervos PR, Plaut GWE. Activity of purified NAD-specific isocitrate dehydrogenase at modulator and substrate concentrations approximating conditions in mitochondria. Metabolism. 1986;35:661–667. - PubMed
-
- Carling D, Mayer FV, Sanders MJ, Gamblin SJ. AMP-activated protein kinase: nature's energy sensor. Nat. Chem. Biol. 2011;7:512–518. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
