

Strategies to discover the structural components of cyst and oocyst walls
- PMID: 24096907
- PMCID: PMC3889564
- DOI: 10.1128/EC.00213-13
Strategies to discover the structural components of cyst and oocyst walls
Abstract
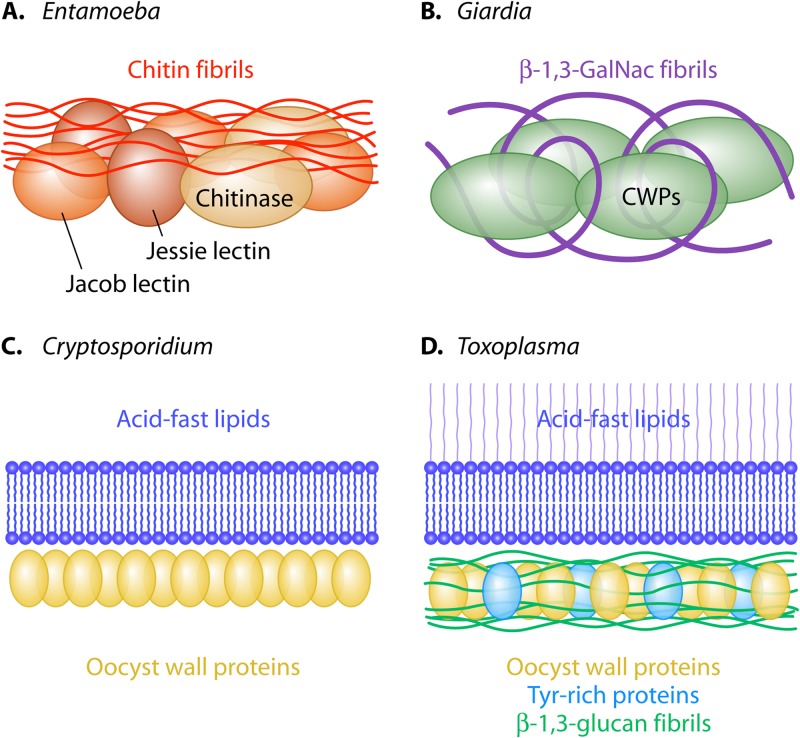
Cysts of Giardia lamblia and Entamoeba histolytica and oocysts of Toxoplasma gondii and Cryptosporidium parvum are the infectious and sometimes diagnostic forms of these parasites. To discover the structural components of cyst and oocyst walls, we have developed strategies based upon a few simple assumptions. Briefly, the most abundant wall proteins are identified by monoclonal antibodies or mass spectrometry. Structural components include a sugar polysaccharide (chitin for Entamoeba, β-1,3-linked glucose for Toxoplasma, and β-1,3-linked GalNAc for Giardia) and/or acid-fast lipids (Toxoplasma and Cryptosporidium). Because Entamoeba cysts and Toxoplasma oocysts are difficult to obtain, studies of walls of nonhuman pathogens (E. invadens and Eimeria, respectively) accelerate discovery. Biochemical methods to dissect fungal walls work well for cyst and oocyst walls, although the results are often unexpected. For example, echinocandins, which inhibit glucan synthases and kill fungi, arrest the development of oocyst walls and block their release into the intestinal lumen. Candida walls are coated with mannans, while Entamoeba cysts are coated in a dextran-like glucose polymer. Models for cyst and oocyst walls derive from their structural components and organization within the wall. Cyst walls are composed of chitin fibrils and lectins that bind chitin (Entamoeba) or fibrils of the β-1,3-GalNAc polymer and lectins that bind the polymer (Giardia). Oocyst walls of Toxoplasma have two distinct layers that resemble those of fungi (β-1,3-glucan in the inner layer) or mycobacteria (acid-fast lipids in the outer layer). Oocyst walls of Cryptosporidium have a rigid bilayer of acid-fast lipids and inner layer of oocyst wall proteins.
Figures




References
-
- Baldursson S, Karanis P. 2011. Waterborne transmission of protozoan parasites: review of worldwide outbreaks—an update 2004–2010. Water Res. 45:6603–6614 - PubMed
-
- Ximénez C, Morán P, Rojas L, Valadez A, Gómez A. 2009. Reassessment of the epidemiology of amebiasis: state of the art. Infect. Genet. Evol. 9:1023–1032 - PubMed
-
- Elmore SA, Jones JL, Conrad PA, Patton S, Lindsay DS, Dubey JP. 2010. Toxoplasma gondii: epidemiology, feline clinical aspects, and prevention. Trends Parasitol. 26:190–196 - PubMed
-
- Kotloff KL, Nataro JP, Blackwelder WC, Nasrin D, Farag TH, Panchalingam S, Wu Y, Sow SO, Sur D, Breiman RF, Faruque AS, Zaidi AK, Saha D, Alonso PL, Tamboura B, Sanogo D, Onwuchekwa U, Manna B, Ramamurthy T, Kanungo S, Ochieng JB, Omore R, Oundo JO, Hossain A, Das SK, Ahmed S, Qureshi S, Quadri F, Adegbola RA, Antonio M, Hossain MJ, Akinsola A, Mandomando I, Nhampossa T, Acacio S, Biswas K, O'Reilly CE, Mintz ED, Berkeley LY, Muhsen K, Sommerfelt H, Robins-Browne RM, Levine MM. 2013. Burden and aetiology of diarrhoeal disease in infants and young children in developing countries (the Global Enteric Multicenter Study, GEMS): a prospective, case-control study. Lancet 382:209–222 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
