

Non-epithelial stem cells and cortical interneuron production in the human ganglionic eminences
- PMID: 24097039
- PMCID: PMC4191718
- DOI: 10.1038/nn.3541
Non-epithelial stem cells and cortical interneuron production in the human ganglionic eminences
Abstract
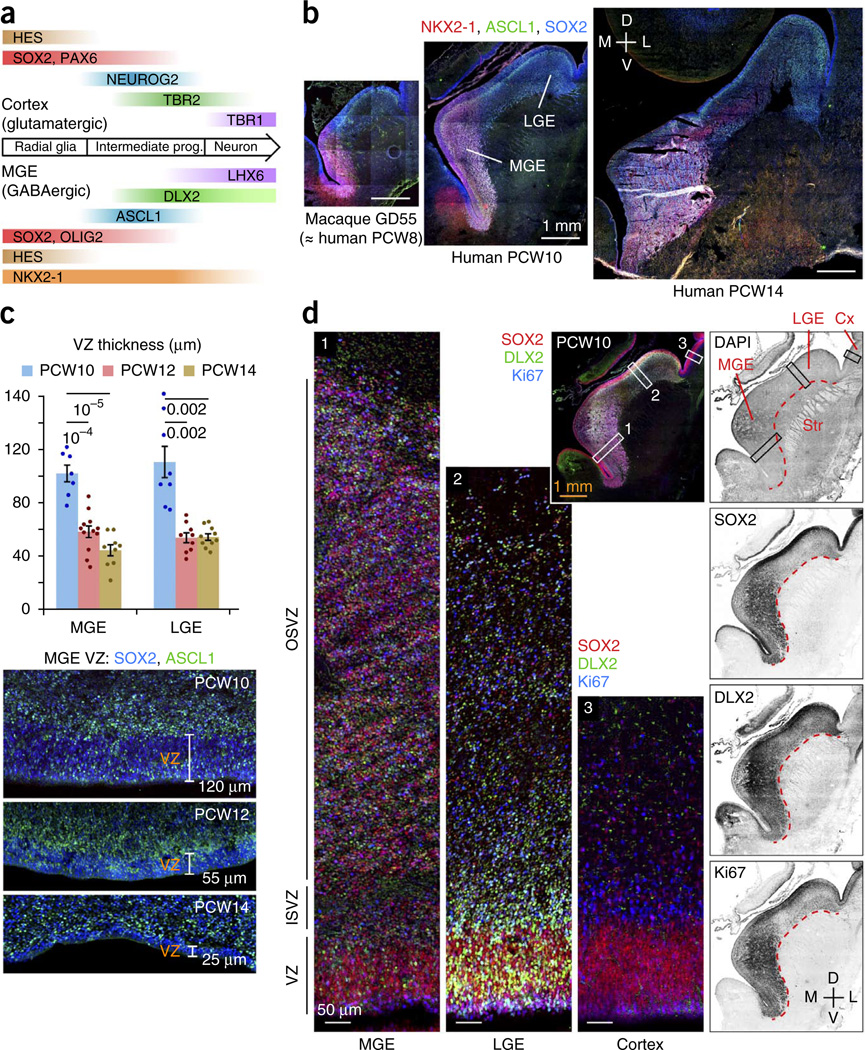
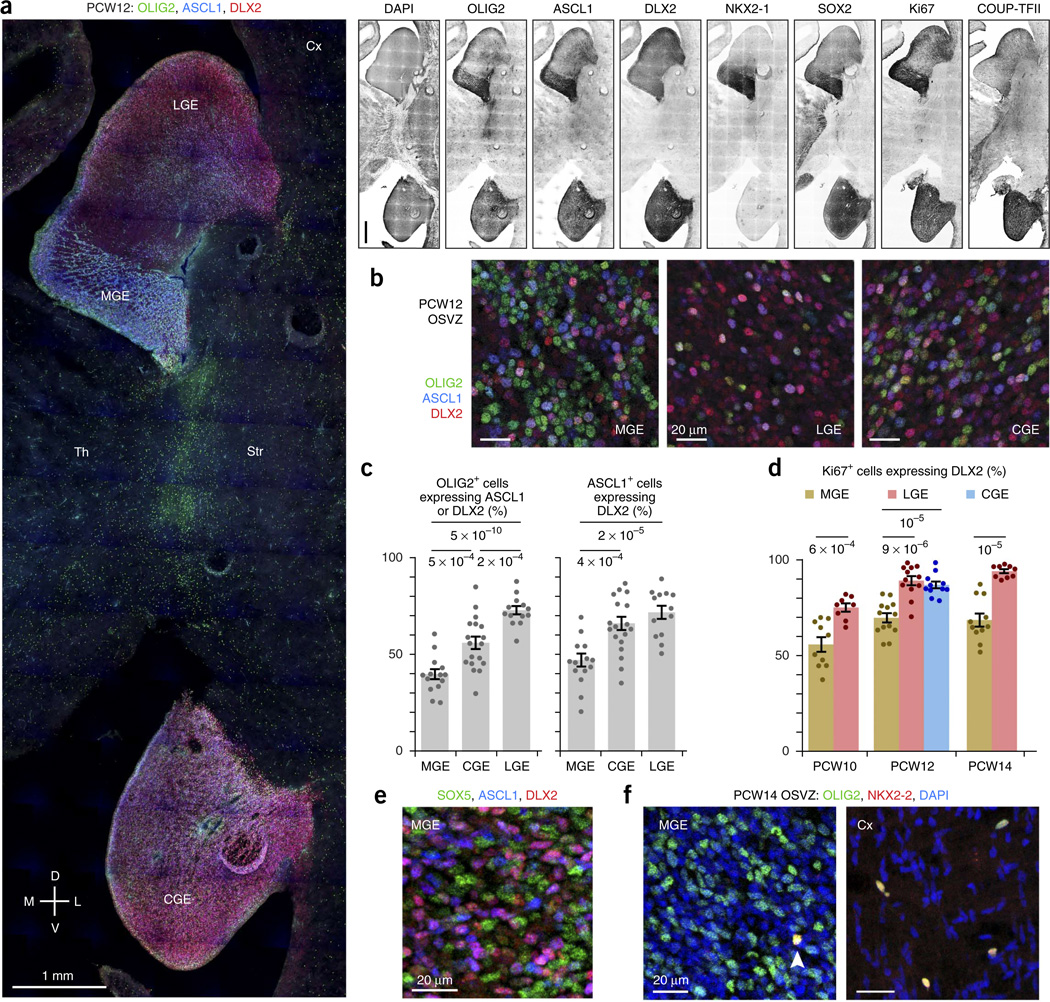
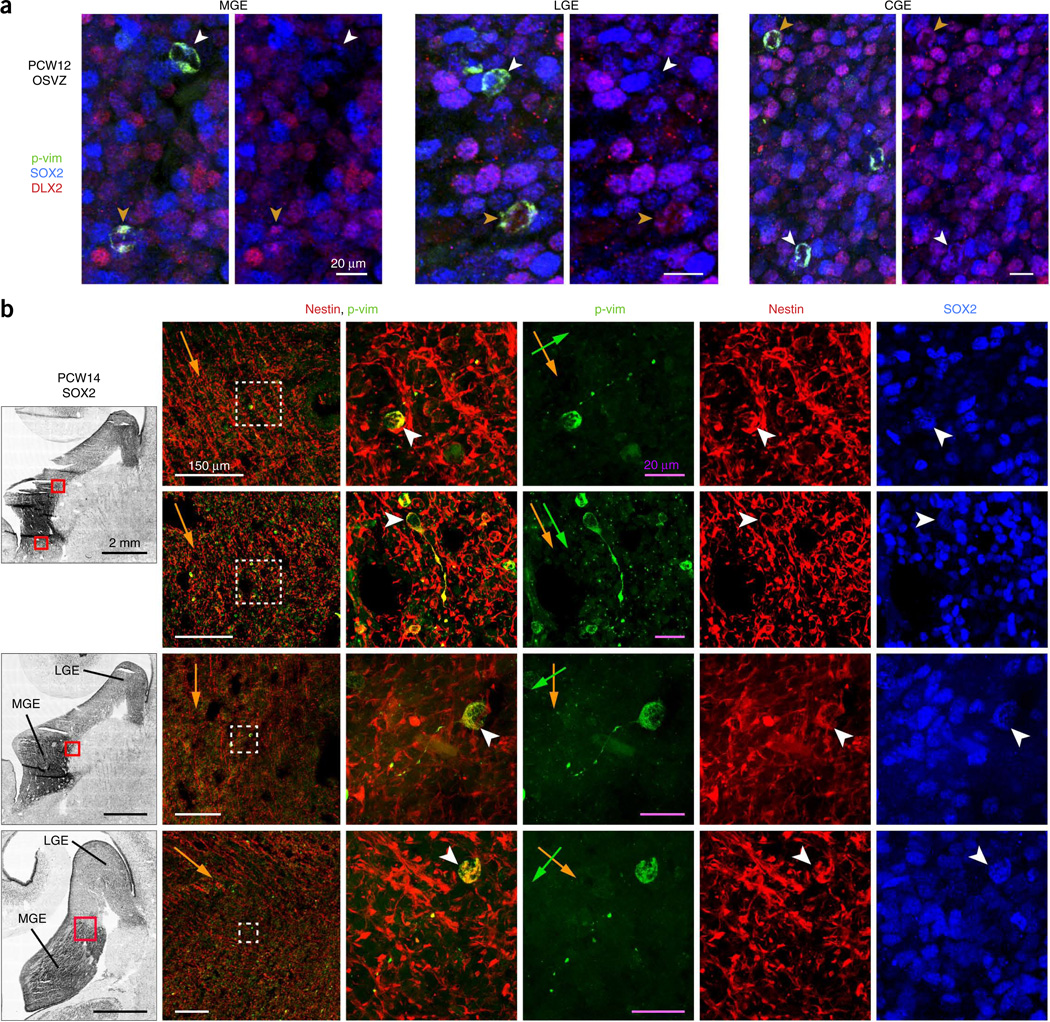
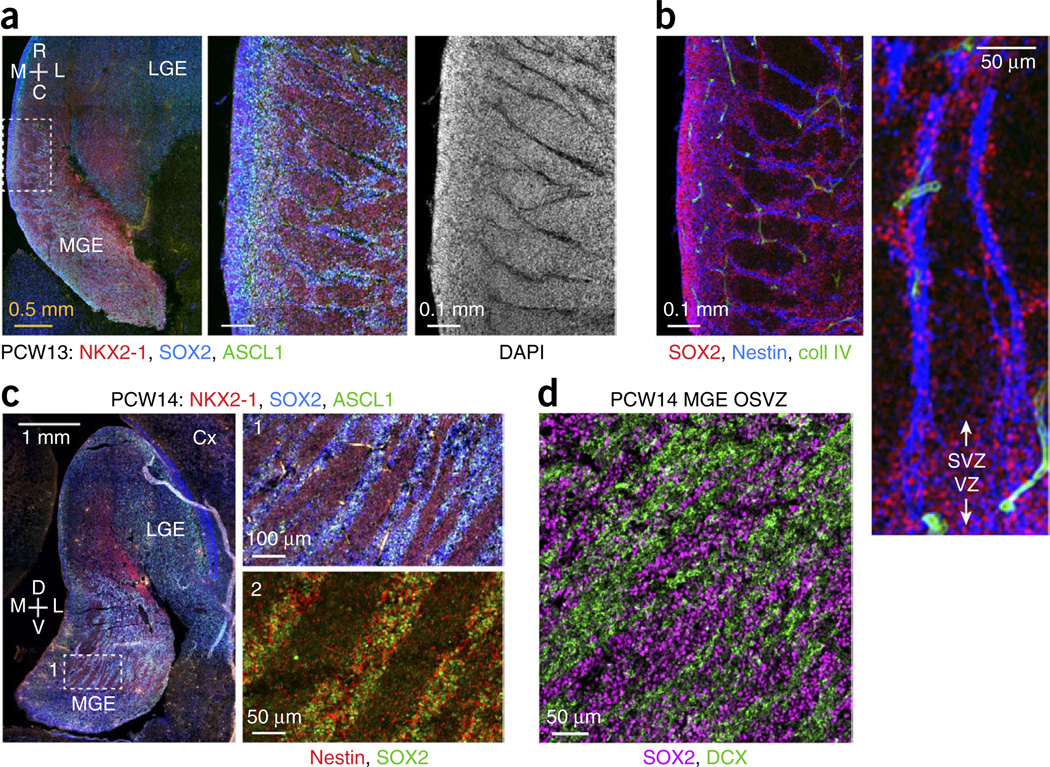
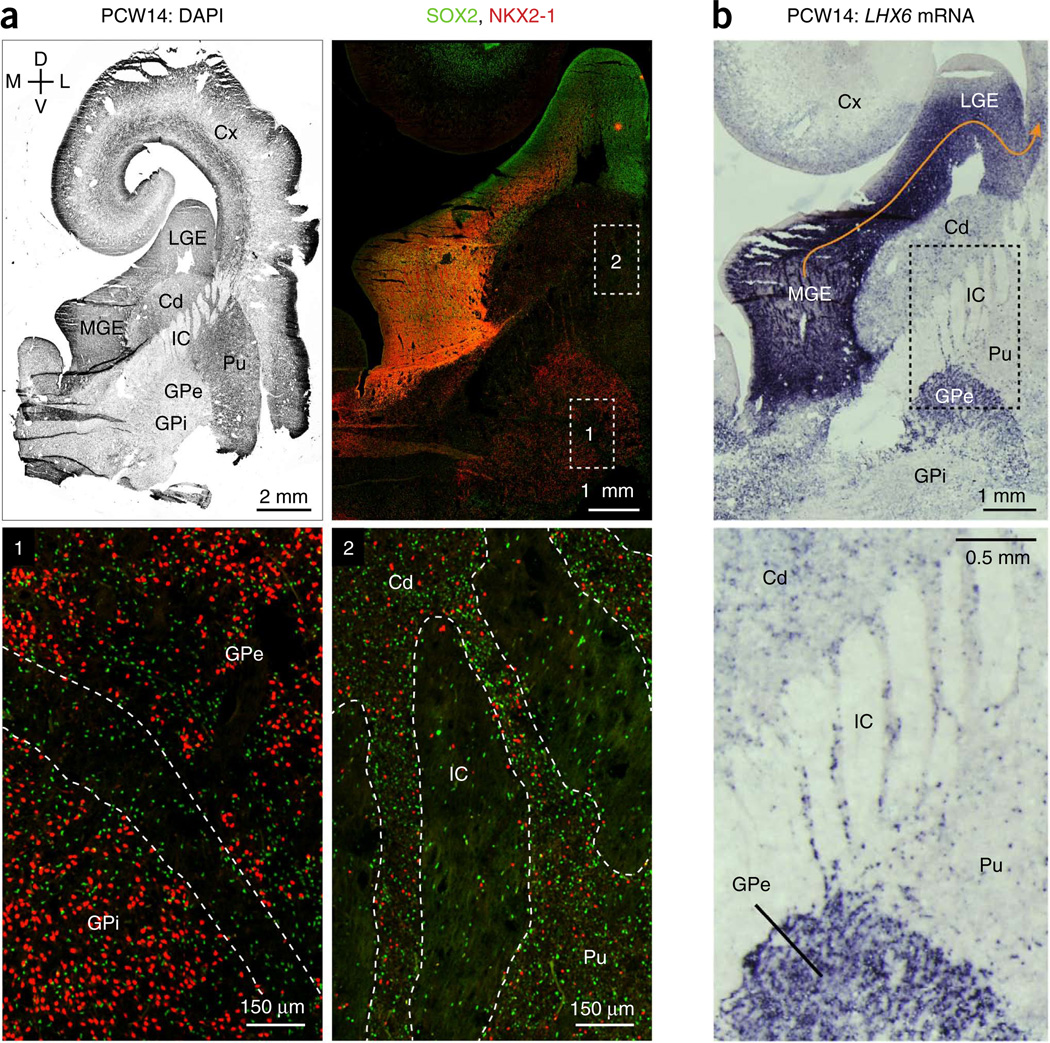
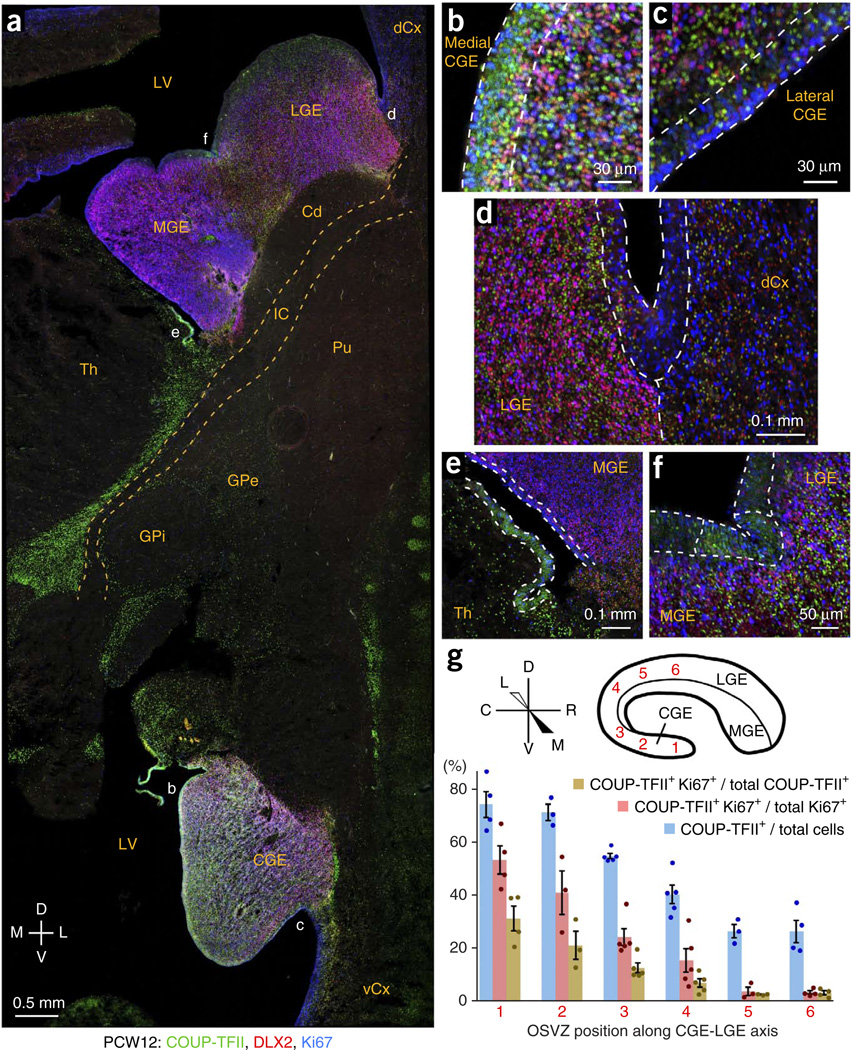
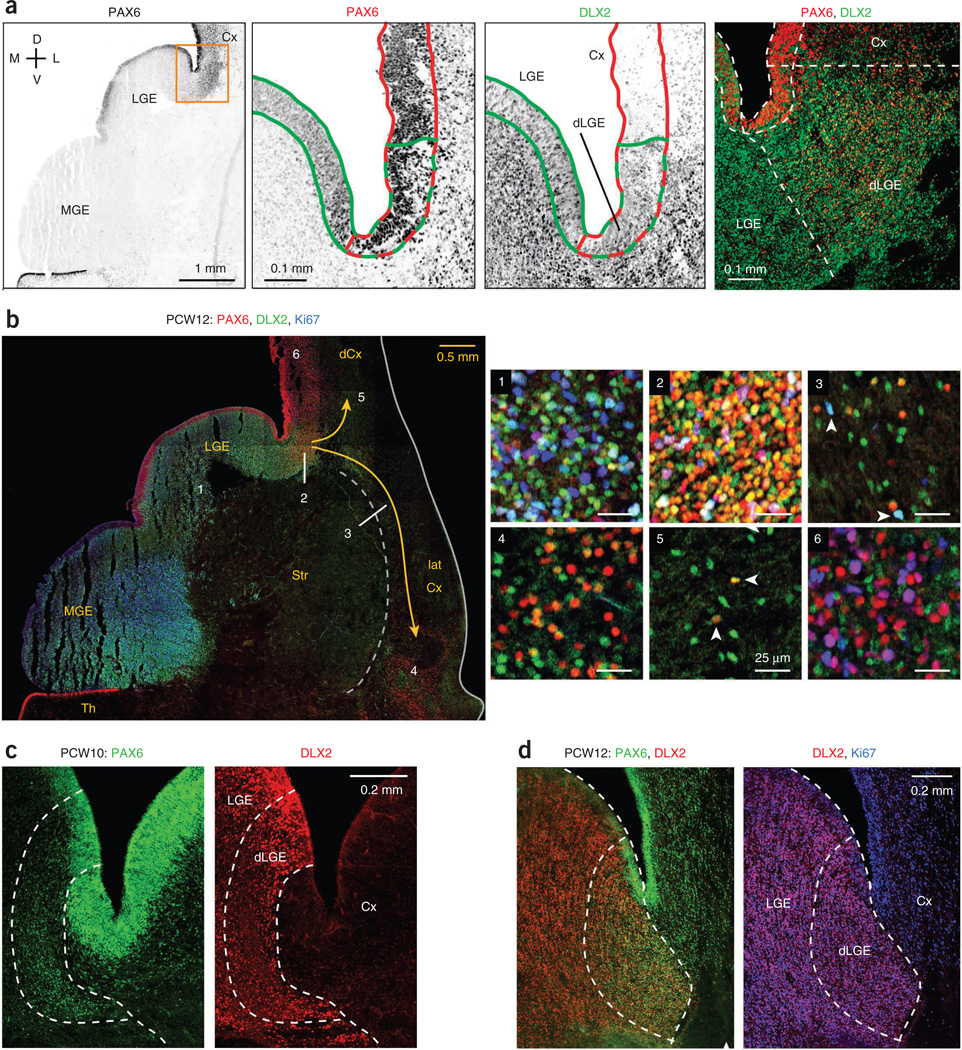
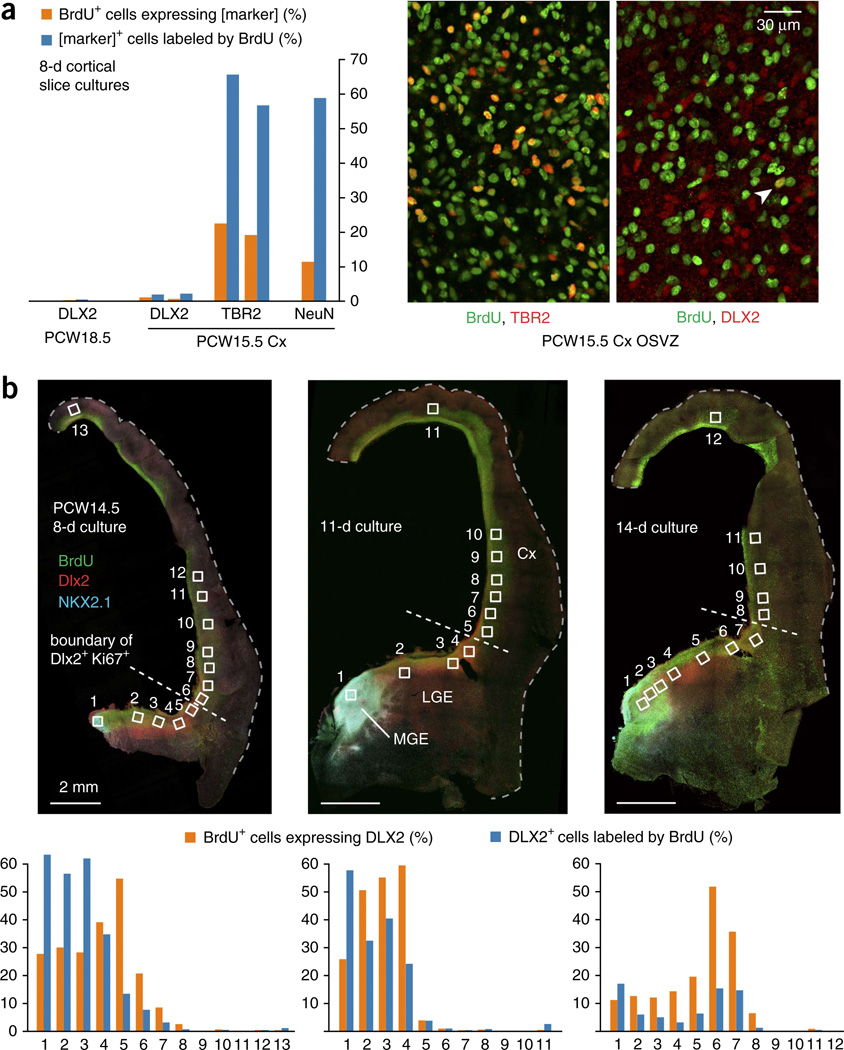
GABAergic cortical interneurons underlie the complexity of neural circuits and are particularly numerous and diverse in humans. In rodents, cortical interneurons originate in the subpallial ganglionic eminences, but their developmental origins in humans are controversial. We characterized the developing human ganglionic eminences and found that the subventricular zone (SVZ) expanded massively during the early second trimester, becoming densely populated with neural stem cells and intermediate progenitor cells. In contrast with the cortex, most stem cells in the ganglionic eminence SVZ did not maintain radial fibers or orientation. The medial ganglionic eminence exhibited unique patterns of progenitor cell organization and clustering, and markers revealed that the caudal ganglionic eminence generated a greater proportion of cortical interneurons in humans than in rodents. On the basis of labeling of newborn neurons in slice culture and mapping of proliferating interneuron progenitors, we conclude that the vast majority of human cortical interneurons are produced in the ganglionic eminences, including an enormous contribution from non-epithelial SVZ stem cells.
Figures
Comment in
-
Neural development: Tracing interneuron roots.Nat Rev Neurosci. 2013 Dec;14(12):818-9. doi: 10.1038/nrn3628. Epub 2013 Oct 23. Nat Rev Neurosci. 2013. PMID: 24149184 No abstract available.
-
Best-laid schemes for interneuron origin of mice and men.Nat Neurosci. 2013 Nov;16(11):1512-4. doi: 10.1038/nn.3557. Nat Neurosci. 2013. PMID: 24165675 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
