

Origin and effect of phototransduction noise in primate cone photoreceptors
- PMID: 24097042
- PMCID: PMC3815624
- DOI: 10.1038/nn.3534
Origin and effect of phototransduction noise in primate cone photoreceptors
Abstract
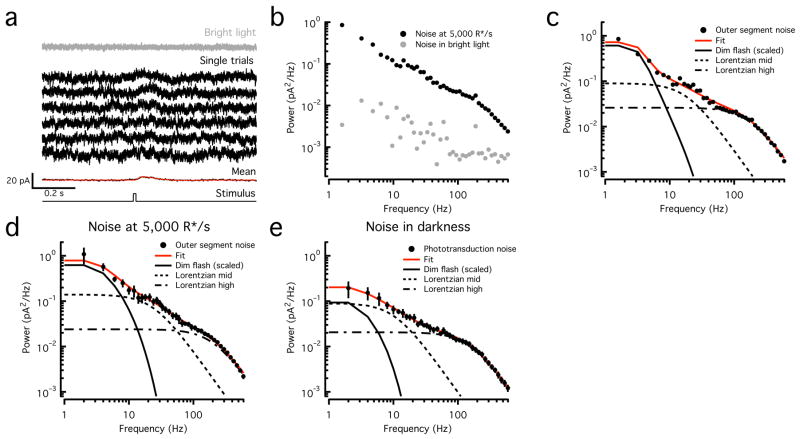
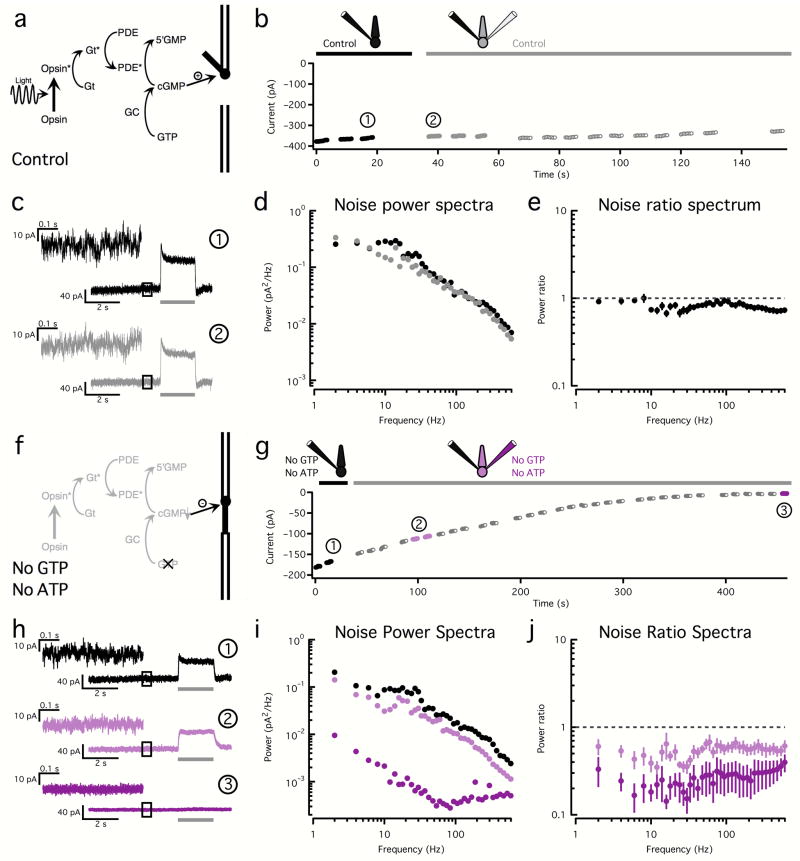
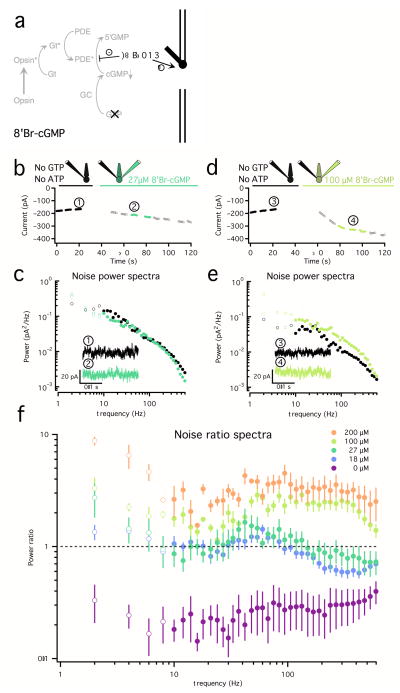
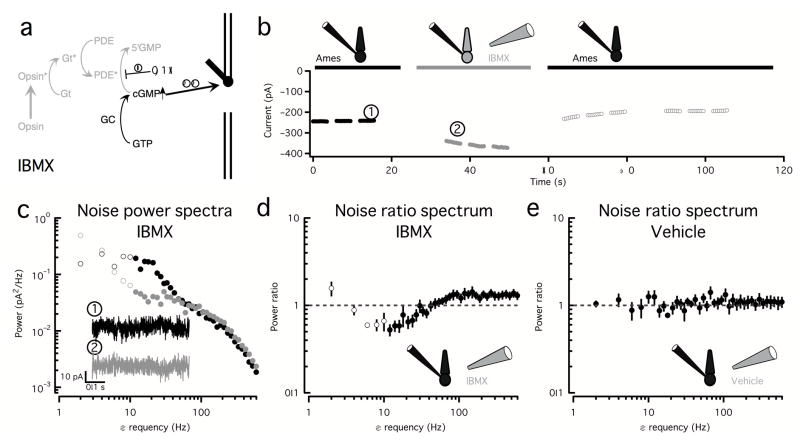
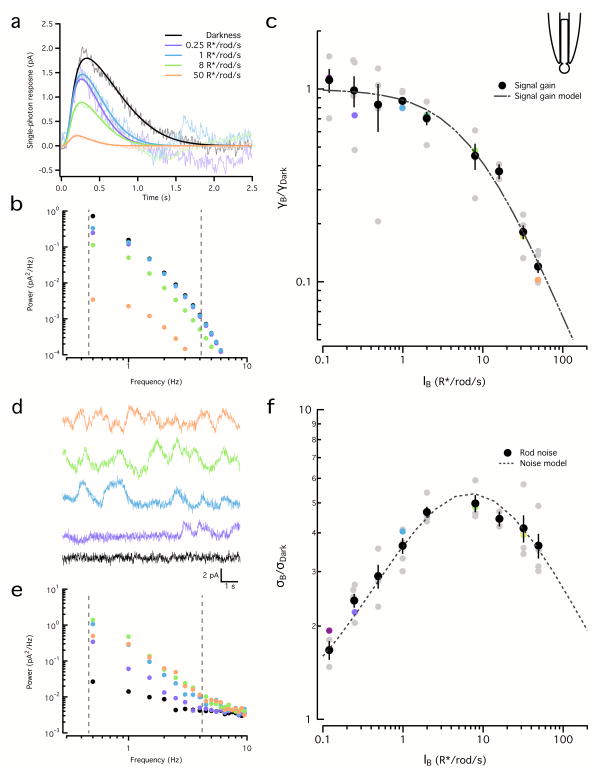
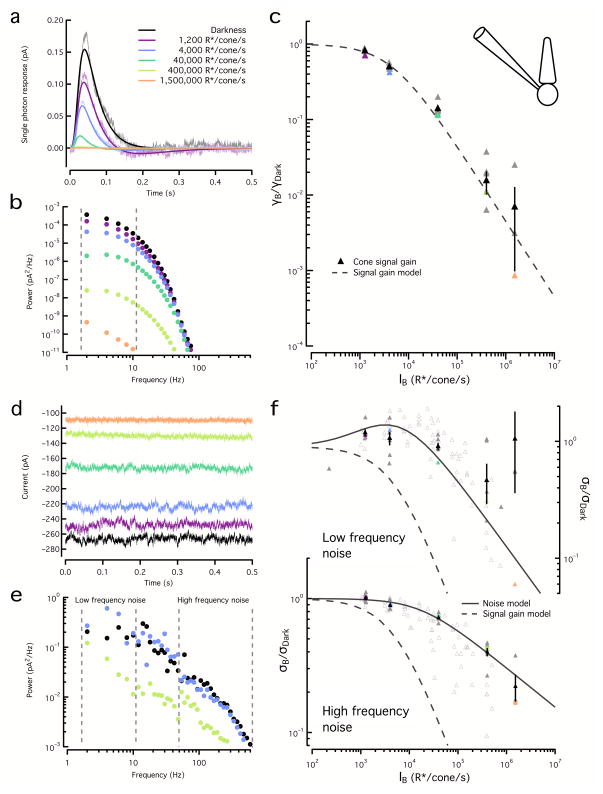
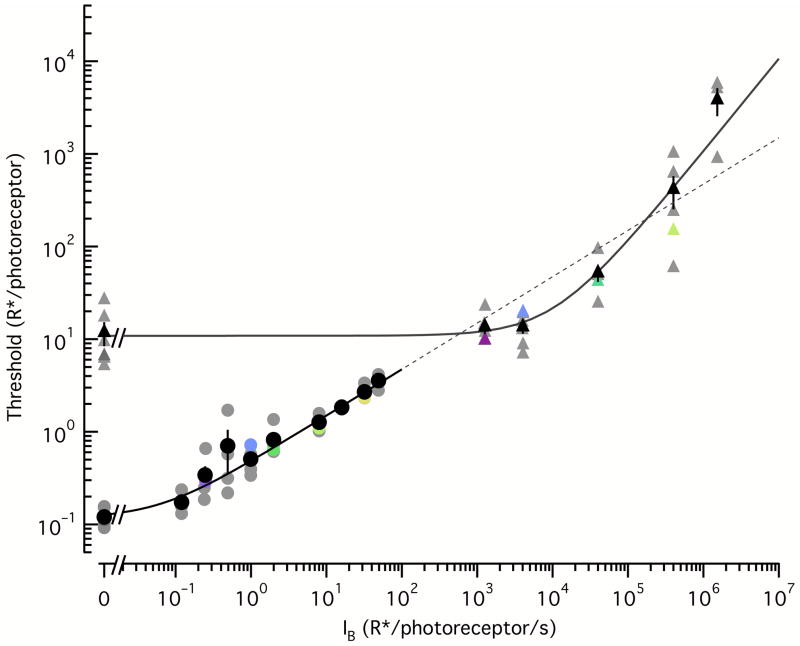
Noise in the responses of cone photoreceptors sets a fundamental limit on visual sensitivity, yet the origin of noise in mammalian cones and its relation to behavioral sensitivity are poorly understood. Our work here on primate cones improves understanding of these issues in three ways. First, we found that cone noise was not dominated by spontaneous photopigment activation or by quantal fluctuations in photon absorption, but was instead dominated by other sources, namely channel noise and fluctuations in cyclic GMP. Second, adaptation in cones, unlike that in rods, affected signal and noise differently. This difference helps to explain why thresholds for rod- and cone-mediated signals have different dependencies on background light level. Third, past estimates of noise in mammalian cones are too high to explain behavioral sensitivity. Our measurements indicate a lower level of cone noise and therefore help to reconcile physiological and behavioral estimates of cone noise and sensitivity.
Figures
References
-
- Donner K. Noise and the absolute thresholds of cone and rod vision. Vision Res. 1992;32:853–866. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
