

Macrophages lift off surface-bound bacteria using a filopodium-lamellipodium hook-and-shovel mechanism
- PMID: 24097079
- PMCID: PMC3791455
- DOI: 10.1038/srep02884
Macrophages lift off surface-bound bacteria using a filopodium-lamellipodium hook-and-shovel mechanism
Abstract
To clear pathogens from host tissues or biomaterial surfaces, phagocytes have to break the adhesive bacteria-substrate interactions. Here we analysed the mechanobiological process that enables macrophages to lift-off and phagocytose surface-bound Escherichia coli (E. coli). In this opsonin-independent process, macrophage filopodia hold on to the E. coli fimbriae long enough to induce a local protrusion of a lamellipodium. Specific contacts between the macrophage and E. coli are formed via the glycoprotein CD48 on filopodia and the adhesin FimH on type 1 fimbriae (hook). We show that bacterial detachment from surfaces occurrs after a lamellipodium has protruded underneath the bacterium (shovel), thereby breaking the multiple bacterium-surface interactions. After lift-off, the bacterium is engulfed by a phagocytic cup. Force activated catch bonds enable the long-term survival of the filopodium-fimbrium interactions while soluble mannose inhibitors and CD48 antibodies suppress the contact formation and thereby inhibit subsequent E. coli phagocytosis.
Figures

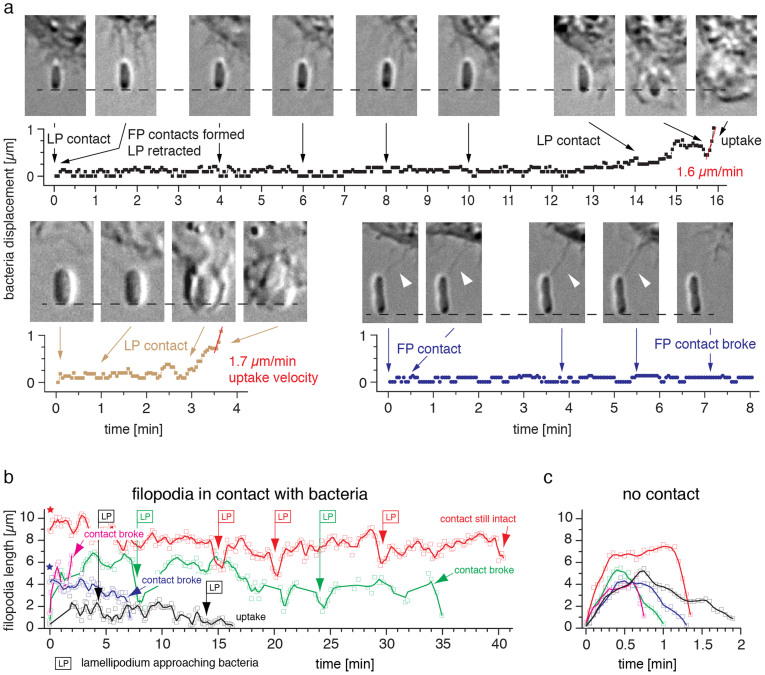
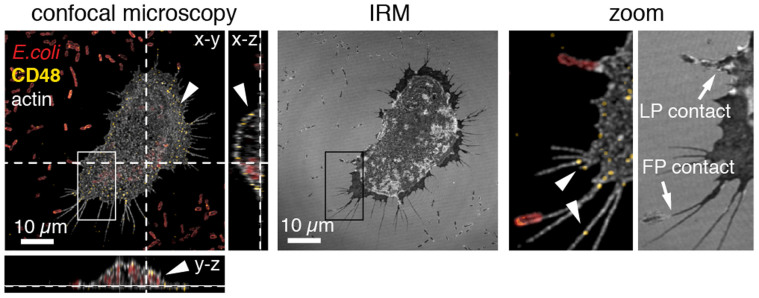
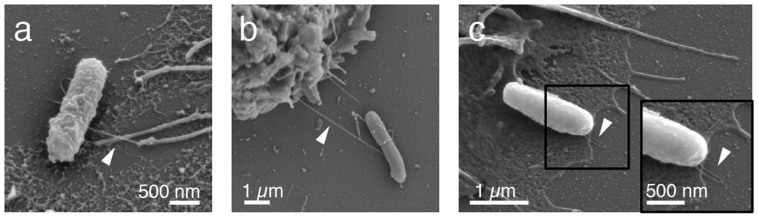


 ), or the non-fimbriated E. coli parent strain AAEC191A was used (Δfim,
), or the non-fimbriated E. coli parent strain AAEC191A was used (Δfim,  ). (b) Fraction of E. coli FimH-j96 that adhered firmly on CD48, 1 M and 3 M. Bacteria were defined firmly adhering if they moved less than one-half of a bacterial diameter over >30 s. (c) Effect of different FimH variants on bacterial accumulation to 1 M and CD48. While the low binding FimH-f18 strain only adhered to CD48 and 1 M above a critical shear stress, FimH-j96 E. coli accumulated on CD48 without any shear threshold.
). (b) Fraction of E. coli FimH-j96 that adhered firmly on CD48, 1 M and 3 M. Bacteria were defined firmly adhering if they moved less than one-half of a bacterial diameter over >30 s. (c) Effect of different FimH variants on bacterial accumulation to 1 M and CD48. While the low binding FimH-f18 strain only adhered to CD48 and 1 M above a critical shear stress, FimH-j96 E. coli accumulated on CD48 without any shear threshold.
References
-
- Aderem A. Phagocytosis and the inflammatory response. J. Infect. Dis. 187 Suppl 2, S340–345 (2003). - PubMed
-
- Mahdavian Delavary B., van der Veer W. M., van Egmond M., Niessen F. B. & Beelen R. H. J. Macrophages in skin injury and repair. Immunobiology 216, 753–762 (2011). - PubMed
-
- Anderson J. M. Biological responses to materials. Annu. Rev. Mater Res. 31, 81–110 (2001).
-
- Nielubowicz G. R. & Mobley H. L. T. Host-pathogen interactions in urinary tract infection. Nat. Rev. Urol. 7, 430–441 (2010). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
