

Review
doi: 10.1101/cshperspect.a012740.
Repair of strand breaks by homologous recombination
Affiliations
- PMID: 24097900
- PMCID: PMC3809576
- DOI: 10.1101/cshperspect.a012740
Item in Clipboard
Review
Repair of strand breaks by homologous recombination
Cold Spring Harb Perspect Biol.
.
Abstract
In this review, we discuss the repair of DNA double-strand breaks (DSBs) using a homologous DNA sequence (i.e., homologous recombination [HR]), focusing mainly on yeast and mammals. We provide a historical context for the current view of HR and describe how DSBs are processed during HR as well as interactions with other DSB repair pathways. We discuss the enzymology of the process, followed by studies on DSB repair in living cells. Whenever possible, we cite both original articles and reviews to aid the reader for further studies.
Figures
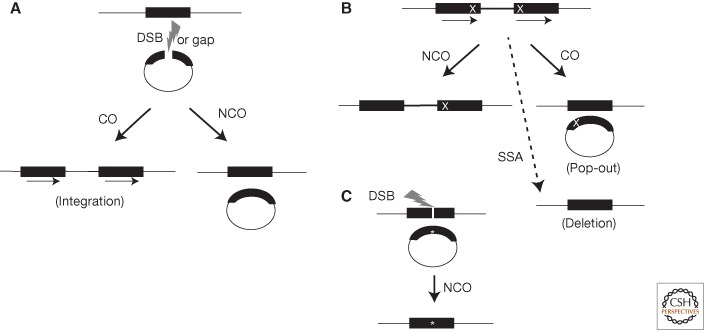
HR events. (A) Plasmid-by-chromosome HR initiated by a DSB or gap (lightning bolt) in the plasmid. The black bars represent the homology between the chromosome and plasmid. In this case, the chromosomal sequence is used as the donor template to repair the gap in the recipient plasmid. Crossovers (COs) lead to plasmid integration and the formation of direct repeats (indicated by arrows). Noncrossovers (NCOs) are also detected if the plasmid contains an origin of replication. (B) Recombination between direct repeats is frequently used to assay HR. Different mutations (X) are present in each repeat. HR associated with gene conversion of one mutation leads to restoration of one intact repeat (solid black bars), which is wild type. NCOs maintain the direct repeat configuration, whereas a CO leads to a plasmid “pop-out” event. Whereas bona fide HR events (i.e., those involving gene conversion) maintain both repeats, SSA (for details, see Fig. 2) leads to deletion of one repeat and the segment between the repeats. (C) DSB-induced gene targeting. Because nonhomologous integration of plasmids in mammalian cells is so efficient, creation of a site-specific DSB in the chromosome is used to induce HR. For the purposes of genome modification (*), the repair of this DSB can be from an incoming plasmid donor sequence, which can be circular (shown) or linear (not shown).
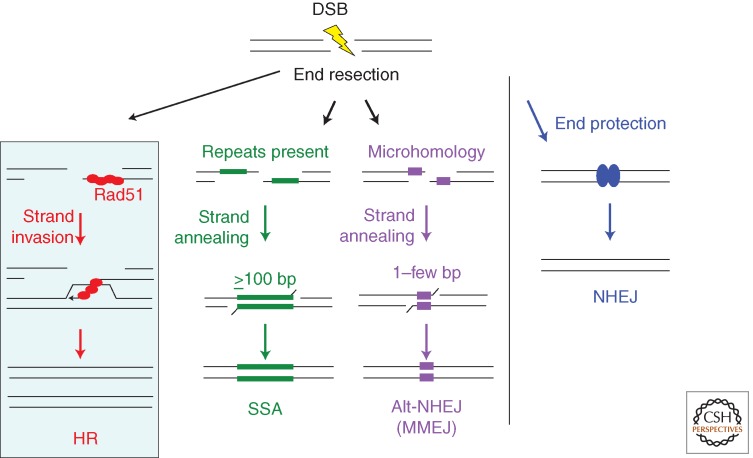
HR is only one pathway of DSB repair and can collaborate with and compete with other pathways to repair DSBs. An early determinant of DSB repair pathway choice is DNA end resection—the processing of DNA ends to generate 3′ single-strands, which is required for HR but inhibits canonical NHEJ in which DNA ends are protected with minimal processing before joining. The defining step of HR is strand invasion by Rad51 or a related recombinase (red balls). Rad51 forms a nucleoprotein filament on single-stranded DNA after end resection. End resection also provides an intermediate in nonconservative single-strand annealing (SSA) and alternative-NHEJ (alt-NHEJ) involving microhomology, as it gives rise to single strands, which can anneal at long (SSA) or short (alt-NHEJ) complementary sequences (green and purple bars, respectively). HR and SSA require more extensive end resection than alt-NHEJ, because minimal end resection will uncover microhomologies present near DNA ends. Alt-NHEJ involving microhomology is also termed microhomology-mediated NHEJ (MMEJ).
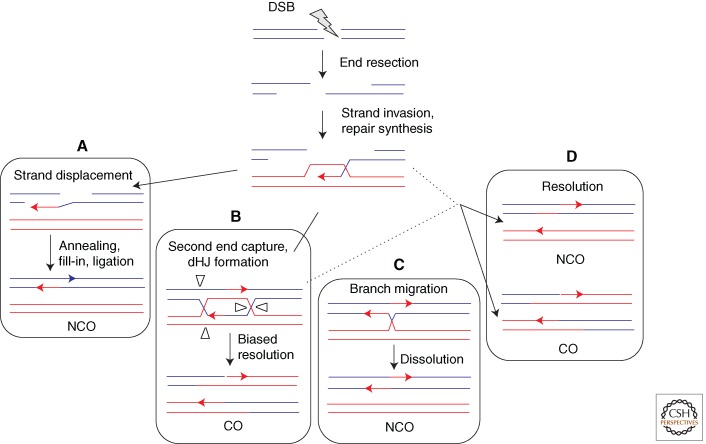
HR of a resected DSB after strand invasion and repair synthesis showing independent and interrelated steps for resolution. (A) Synthesis-dependent strand annealing involves displacement by DNA helicase(s) of the newly synthesized strand. The two ends can anneal by their shared complementarity. Synthesis, nuclease, and ligase reactions finish the event. (B) Double Holliday junctions (dHJs) are formed when the D loop captures the second end. In meiotic cells, dHJ resolution is biased to give COs. (C) dHJs can be dissolved without crossover by the action of the BLM(Sgs1)/TOP3α/RMI1 complex. (D) Resolution of intermediates that escape the BLM(Sgs1) complex may occur from the action of different resolvases (e.g., MUS81/EME1[Mms4], GEN1[Yen1], etc.).
References
-
- Allers T, Lichten M 2001. Differential timing and control of noncrossover and crossover recombination during meiosis. Cell 106: 47–57 - PubMed
-
- Aten JA, Stap J, Krawczyk PM, van Oven CH, Hoebe RA, Essers J, Kanaar R 2004. Dynamics of DNA double-strand breaks revealed by clustering of damaged chromosome domains. Science 303: 92–95 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases