

Heme oxygenase-1 exerts a protective role in ovalbumin-induced neutrophilic airway inflammation by inhibiting Th17 cell-mediated immune response
- PMID: 24097973
- PMCID: PMC3843074
- DOI: 10.1074/jbc.M113.494369
Heme oxygenase-1 exerts a protective role in ovalbumin-induced neutrophilic airway inflammation by inhibiting Th17 cell-mediated immune response
Abstract
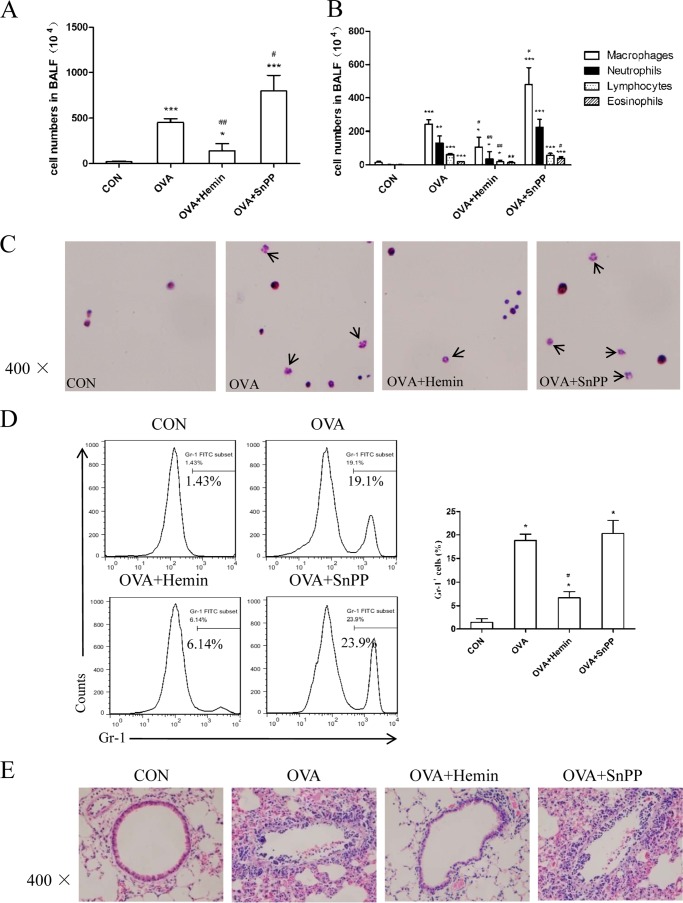
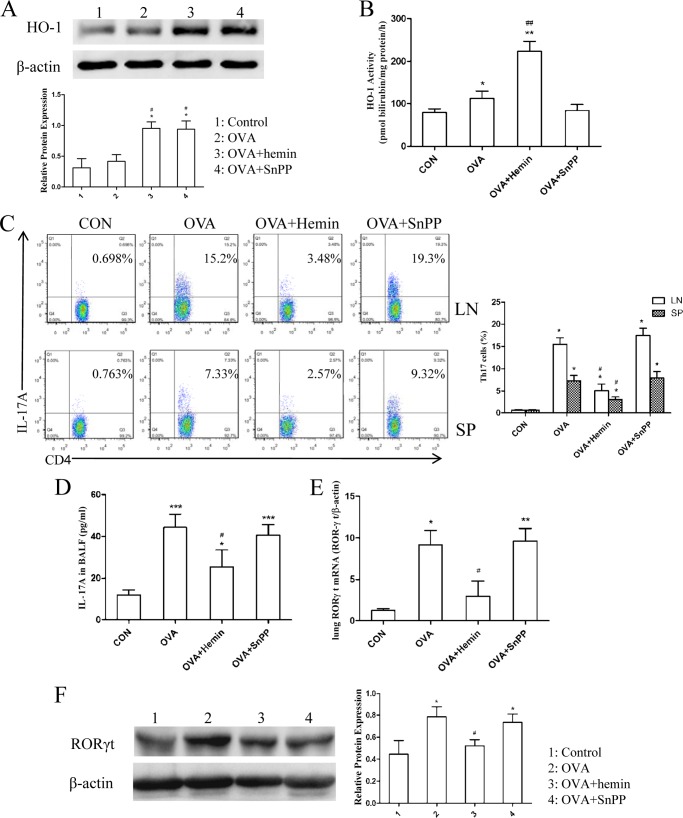
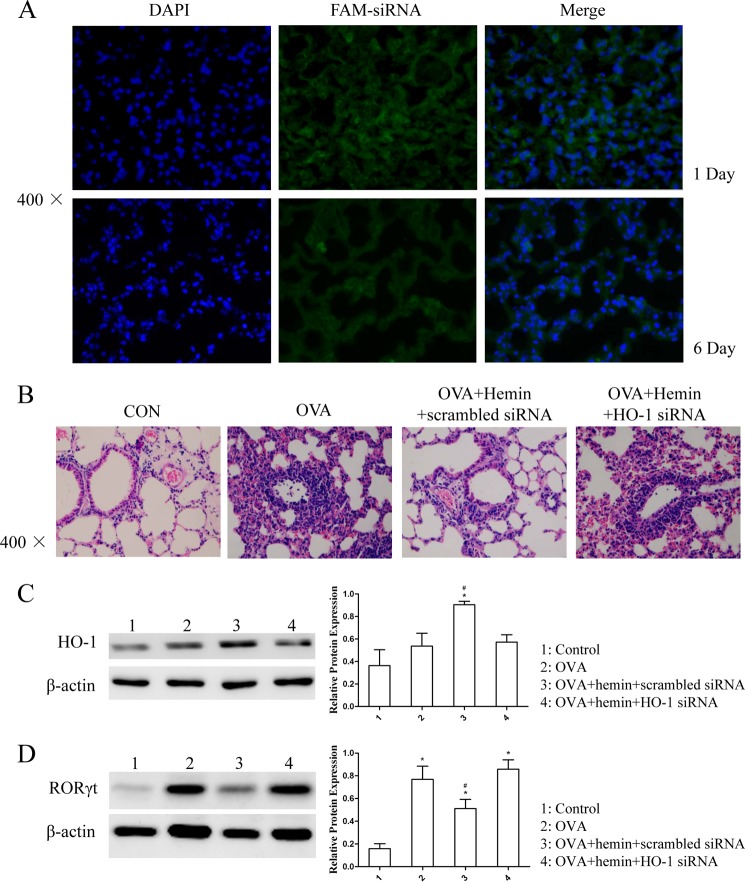
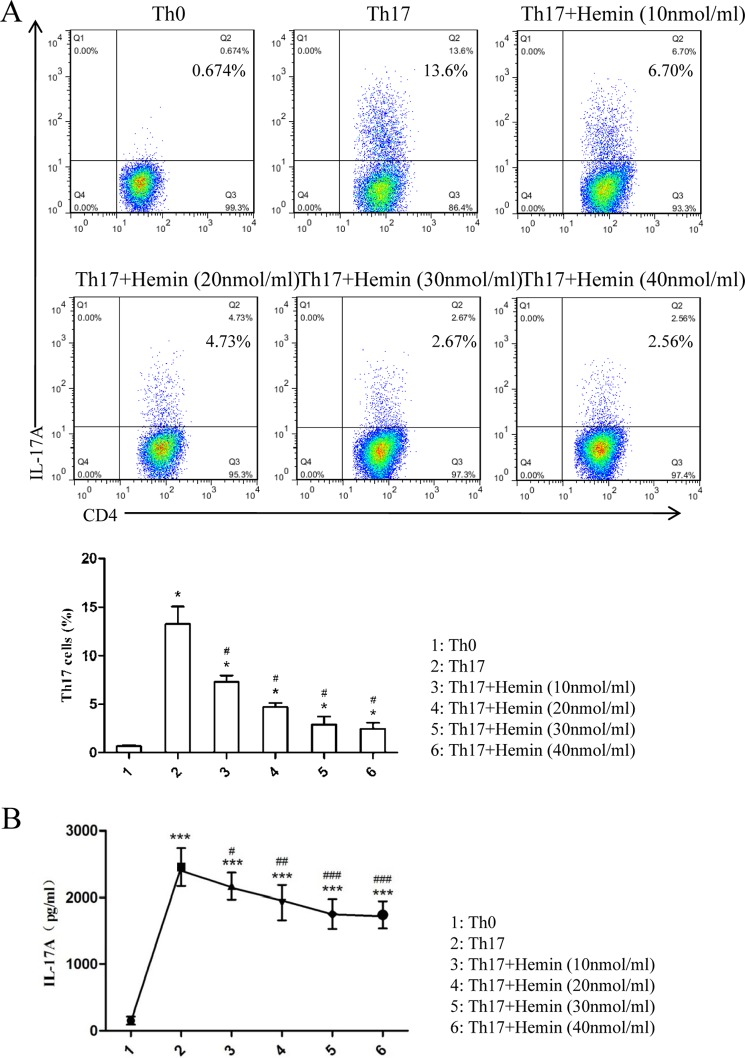
Allergic asthma is conventionally considered as a Th2 immune response characterized by eosinophilic inflammation. Recent investigations revealed that Th17 cells play an important role in the pathogenesis of non-eosinophilic asthma (NEA), resulting in steroid-resistant neutrophilic airway inflammation. Heme oxygenase-1 (HO-1) has anti-inflammation, anti-oxidation, and anti-apoptosis functions. However, its role in NEA is still unclear. Here, we explore the role of HO-1 in a mouse model of NEA. HO-1 inducer hemin or HO-1 inhibitor tin protoporphyrin IX was injected intraperitoneally into ovalbumin-challenged DO11.10 mice. Small interfering RNA (siRNA) was delivered into mice to knock down HO-1 expression. The results show that induction of HO-1 by hemin attenuated airway inflammation and decreased neutrophil infiltration in bronchial alveolar lavage fluid and was accompanied by a lower proportion of Th17 cells in mediastinal lymph nodes and spleen. More importantly, induction of HO-1 down-regulated Th17-related transcription factor retinoic acid-related orphan receptor γt (RORγt) expression and decreased IL-17A levels, all of which correlated with a decrease in phosphorylated STAT3 (p-STAT3) level and inhibition of Th17 cell differentiation. Consistently, the above events could be reversed by tin protoporphyrin IX. Also, HO-1 siRNA transfection abolished the effect of hemin induced HO-1 in vivo. Meanwhile, the hemin treatment promoted the level of Foxp3 expression and enhanced the proportion of regulatory T cells (Tregs). Collectively, our findings indicate that HO-1 exhibits anti-inflammatory activity in the mouse model of NEA via inhibition of the p-STAT3-RORγt pathway, regulating kinetics of RORγt and Foxp3 expression, thus providing a possible novel therapeutic target in asthmatic patients.
Keywords: Asthma; Heme Oxygenase; Immunology; Inflammation; T Cell; Th17; Treg.
Figures
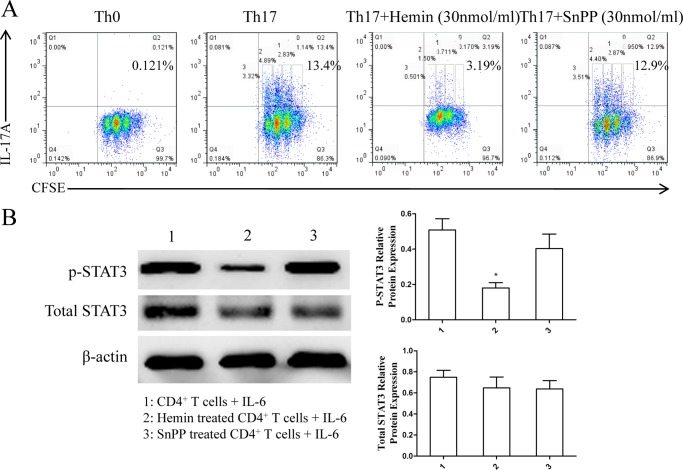
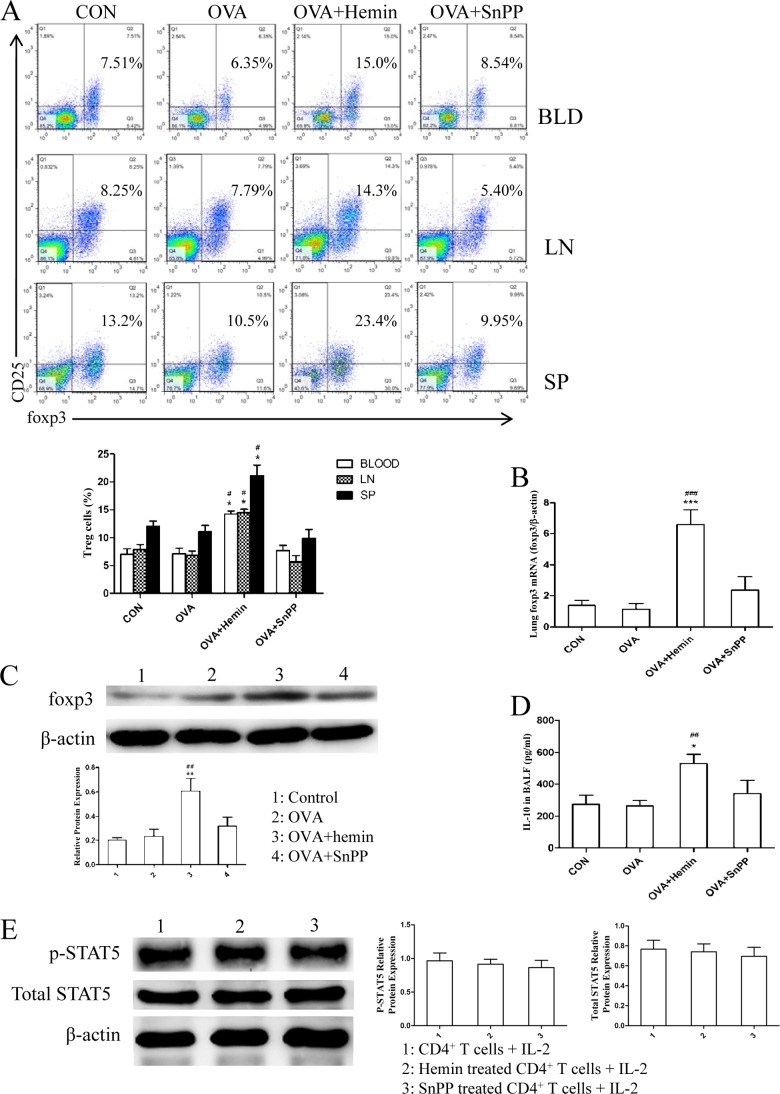
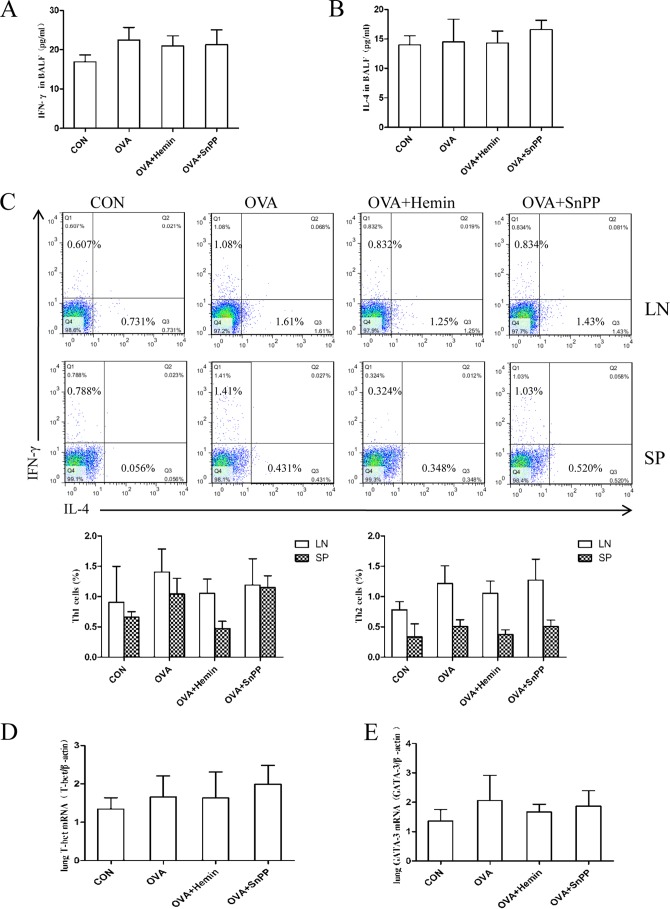
References
-
- Simpson J. L., Scott R., Boyle M. J., Gibson P. G. (2006) Inflammatory subtypes in asthma. Assessment and identification using induced sputum. Respirology 11, 54–61 - PubMed
-
- Kikuchi S., Nagata M., Kikuchi I., Hagiwara K., Kanazawa M. (2005) Association between neutrophilic and eosinophilic inflammation in patients with severe persistent asthma. Int. Arch. Allergy Immunol. 137, 7–11 - PubMed
-
- The ENFUMOSA Study Group (2003) The ENFUMOSA cross-sectional European multicentre study of the clinical phenotype of chronic severe asthma. Eur. Respir. J. 22, 470–477 - PubMed
-
- Oboki K., Ohno T., Saito H., Nakae S. (2008) Th17 and allergy. Allergol Int. 57, 121–134 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
