

Analysis of hyperekplexia mutations identifies transmembrane domain rearrangements that mediate glycine receptor activation
- PMID: 24097980
- PMCID: PMC3837120
- DOI: 10.1074/jbc.M113.513804
Analysis of hyperekplexia mutations identifies transmembrane domain rearrangements that mediate glycine receptor activation
Abstract
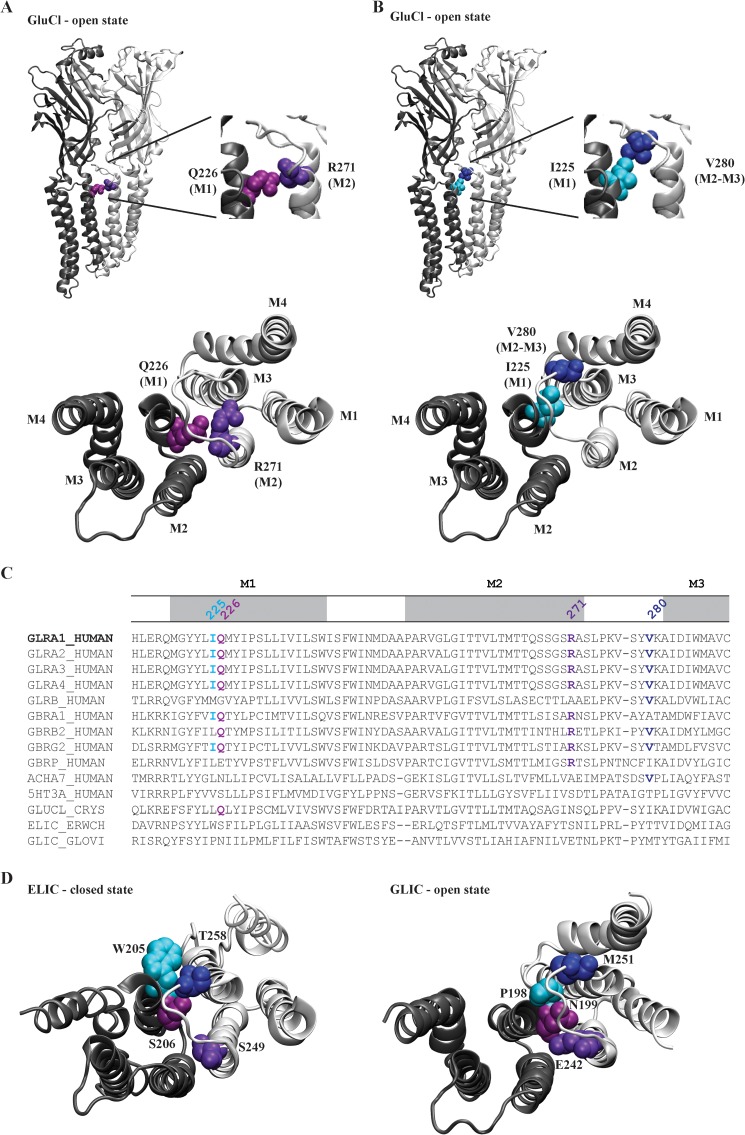
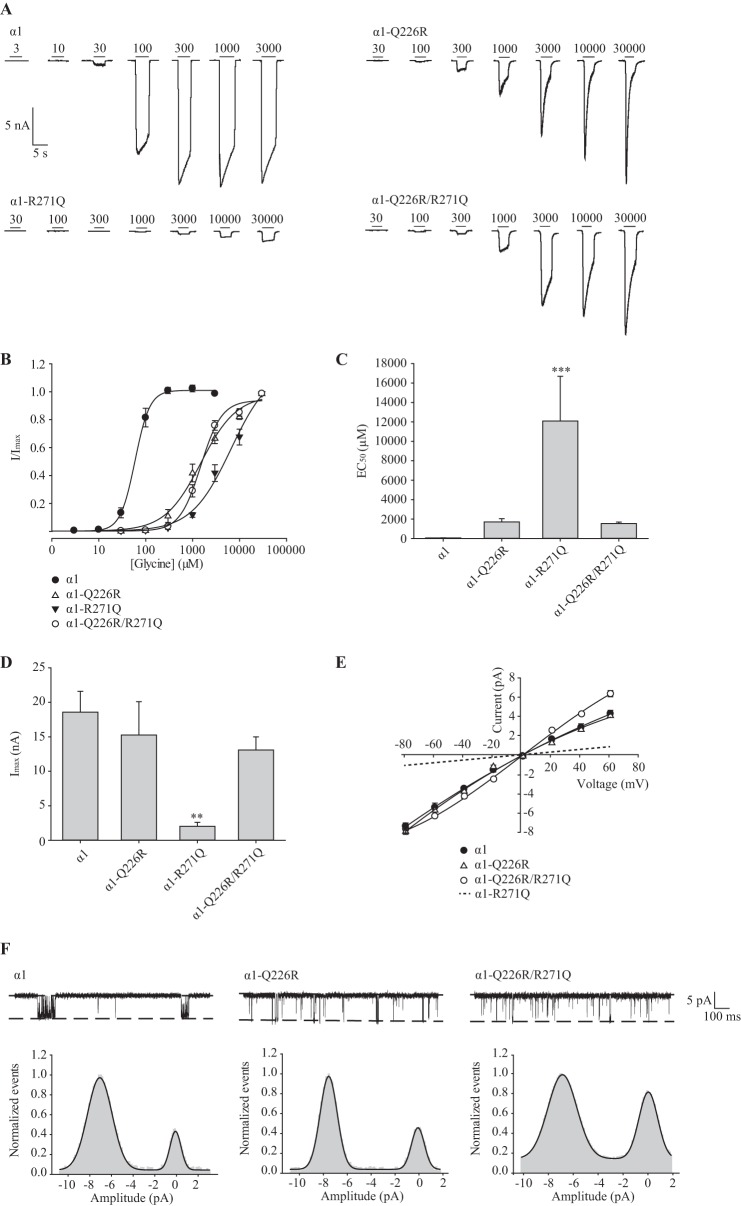
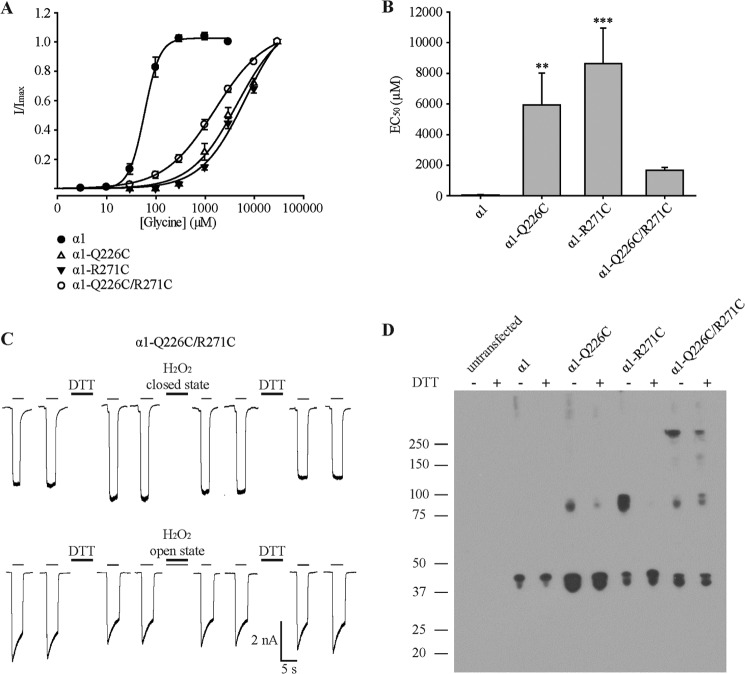
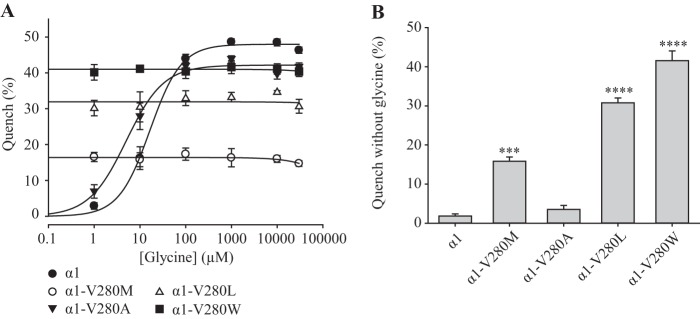
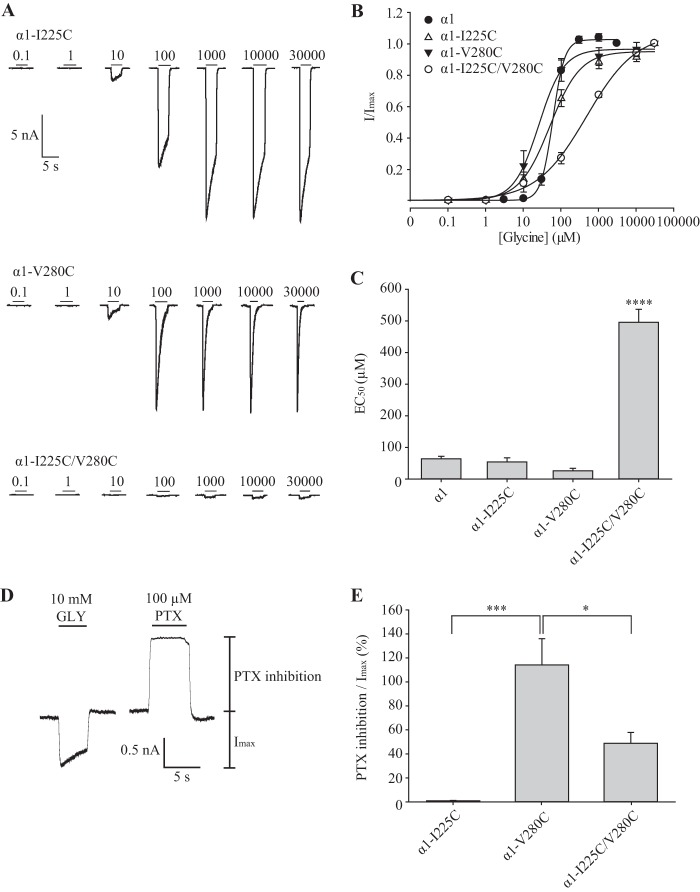
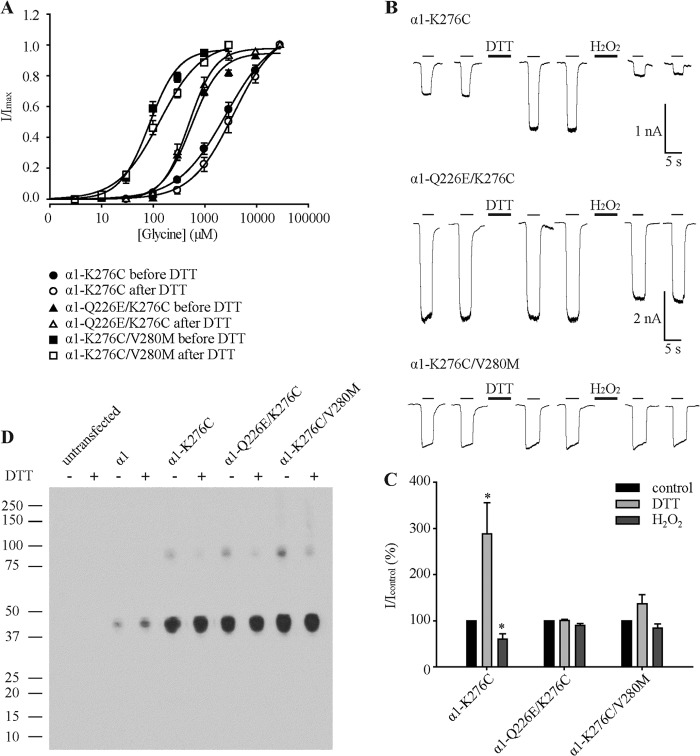
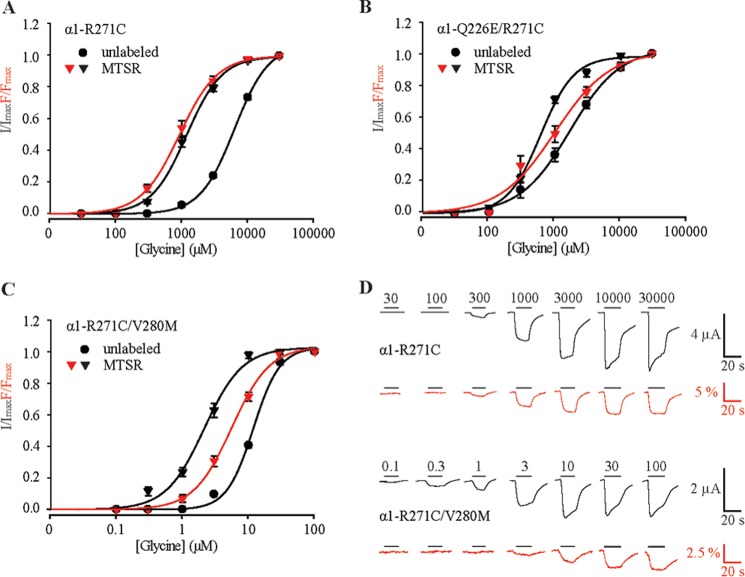
Pentameric ligand-gated ion channels (pLGICs) mediate numerous physiological processes and are therapeutic targets for a wide range of clinical indications. Elucidating the structural differences between their closed and open states may help in designing improved drugs that bias receptors toward the desired conformational state. We recently showed that two new hyperekplexia mutations, Q226E and V280M, induced spontaneous activity in α1 glycine receptors. Gln-226, located near the top of transmembrane (TM) 1, is closely apposed to Arg-271 at the top of TM2 in the neighboring subunit. Using mutant cycle analysis, we inferred that Q226E induces activation via an enhanced electrostatic attraction to Arg-271. This would tilt the top of TM2 toward TM1 and hence away from the pore axis to open the channel. We also concluded that the increased side chain volume of V280M, in the TM2-TM3 loop, exerts a steric repulsion against Ile-225 at the top of TM1 in the neighboring subunit. We infer that this steric repulsion would tilt the top of TM3 radially outwards against the stationary TM1 and thus provide space for TM2 to relax away from the pore axis to create an open channel. Because the transmembrane domain movements inferred from this functional analysis are consistent with the structural differences evident in the x-ray atomic structures of closed and open state bacterial pLGICs, we propyose that the model of pLGIC activation as outlined here may be broadly applicable across the eukaryotic pLGIC receptor family.
Keywords: Cys Loop Receptors; Glycine Receptors; Patch Clamp; Receptor Structure-Function; Site-directed Mutagenesis.
Figures
References
-
- Lynch J. W. (2004) Molecular structure and function of the glycine receptor chloride channel. Physiol. Rev. 84, 1051–1095 - PubMed
-
- Lynch J. W. (2009) Native glycine receptor subtypes and their physiological roles. Neuropharmacology 56, 303–309 - PubMed
-
- Fritschy J. M., Harvey R. J., Schwarz G. (2008) Gephyrin. Where do we stand, where do we go? Trend. Neurosci. 31, 257–264 - PubMed
-
- Specht C. G., Izeddin I., Rodriguez P. C., El Beheiry M., Rostaing P., Darzacq X., Dahan M., Triller A. (2013) Quantitative nanoscopy of inhibitory synapses. Counting gephyrin molecules and receptor binding sites. Neuron 79, 308–321 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
