

A genetic RNAi screen for IP₃/Ca²⁺ coupled GPCRs in Drosophila identifies the PdfR as a regulator of insect flight
- PMID: 24098151
- PMCID: PMC3789835
- DOI: 10.1371/journal.pgen.1003849
A genetic RNAi screen for IP₃/Ca²⁺ coupled GPCRs in Drosophila identifies the PdfR as a regulator of insect flight
Abstract
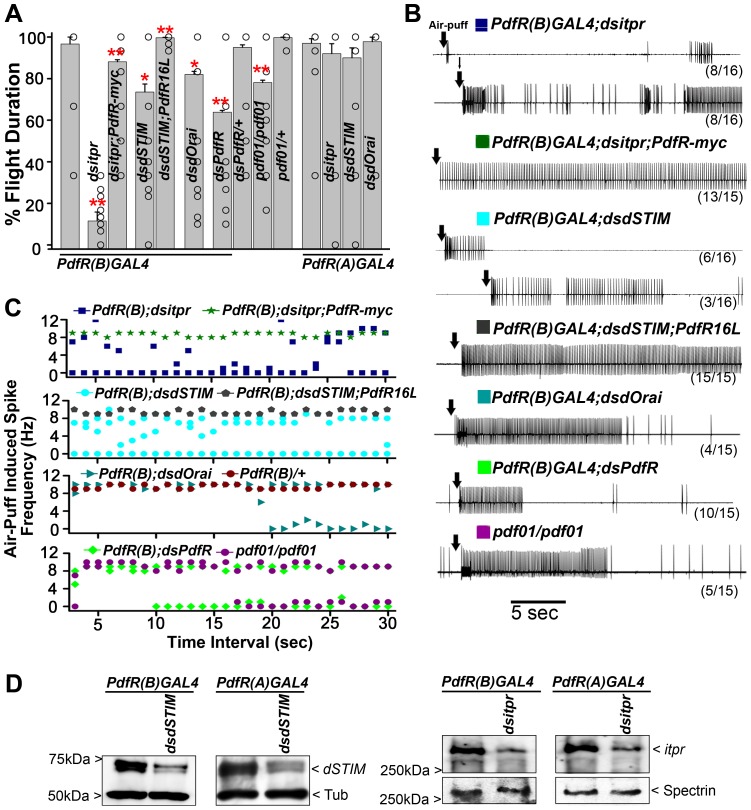
Insect flight is regulated by various sensory inputs and neuromodulatory circuits which function in synchrony to control and fine-tune the final behavioral outcome. The cellular and molecular bases of flight neuromodulatory circuits are not well defined. In Drosophila melanogaster, it is known that neuronal IP3 receptor mediated Ca²⁺ signaling and store-operated Ca²⁺ entry (SOCE) are required for air-puff stimulated adult flight. However, G-protein coupled receptors (GPCRs) that activate intracellular Ca²⁺ signaling in the context of flight are unknown in Drosophila. We performed a genetic RNAi screen to identify GPCRs that regulate flight by activating the IPIP₃ receptor. Among the 108 GPCRs screened, we discovered 5 IPIP₃/Ca²⁺ linked GPCRs that are necessary for maintenance of air-puff stimulated flight. Analysis of their temporal requirement established that while some GPCRs are required only during flight circuit development, others are required both in pupal development as well as during adult flight. Interestingly, our study identified the Pigment Dispersing Factor Receptor (PdfR) as a regulator of flight circuit development and as a modulator of acute flight. From the analysis of PdfR expressing neurons relevant for flight and its well-defined roles in other behavioral paradigms, we propose that PdfR signaling functions systemically to integrate multiple sensory inputs and modulate downstream motor behavior.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures








References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
