

Viral membrane scission
- PMID: 24099087
- PMCID: PMC4286373
- DOI: 10.1146/annurev-cellbio-101011-155838
Viral membrane scission
Abstract
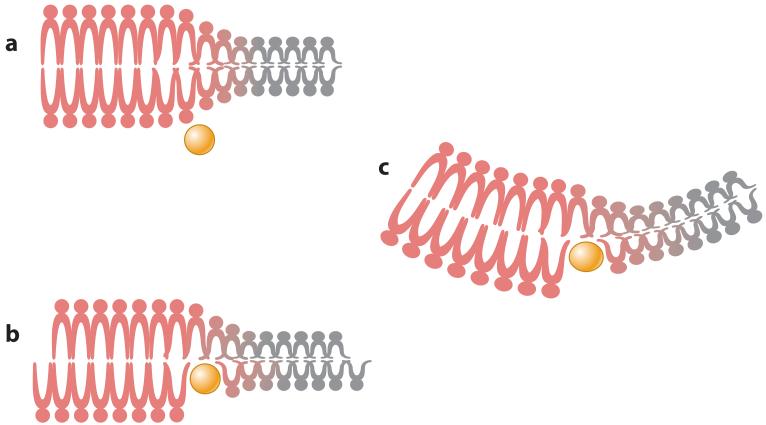
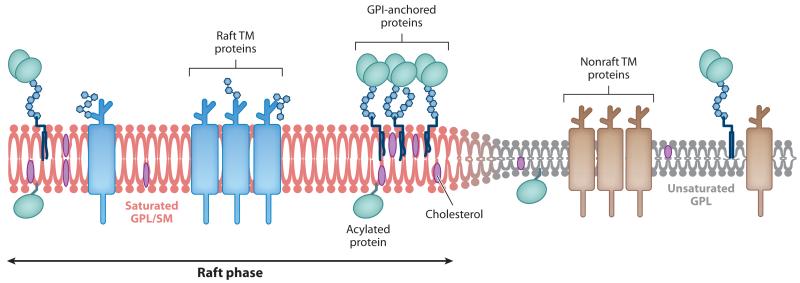
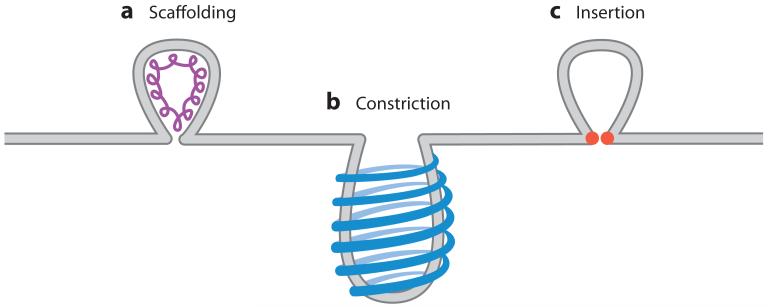
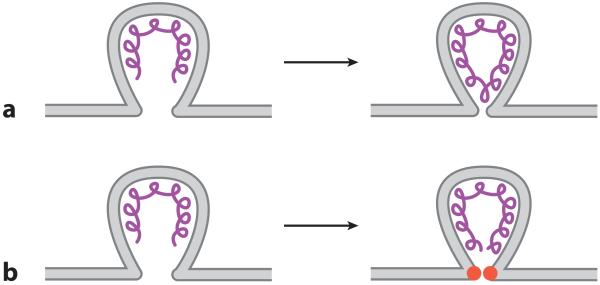
Virus budding is a complex, multistep process in which viral proteins make specific alterations in membrane curvature. Many different viral proteins can deform the membrane and form a budding virion, but very few can mediate membrane scission to complete the budding process. As a result, enveloped viruses have developed numerous ways of facilitating membrane scission, including hijacking host cellular scission machinery and expressing their own scission proteins. These proteins mediate scission in very different ways, though the biophysical mechanics underlying their actions may be similar. In this review, we explore the mechanisms of membrane scission and the ways in which enveloped viruses use these systems to mediate the release of budding virions.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
