

Pin1 promotes histone H1 dephosphorylation and stabilizes its binding to chromatin
- PMID: 24100296
- PMCID: PMC3798258
- DOI: 10.1083/jcb.201305159
Pin1 promotes histone H1 dephosphorylation and stabilizes its binding to chromatin
Abstract
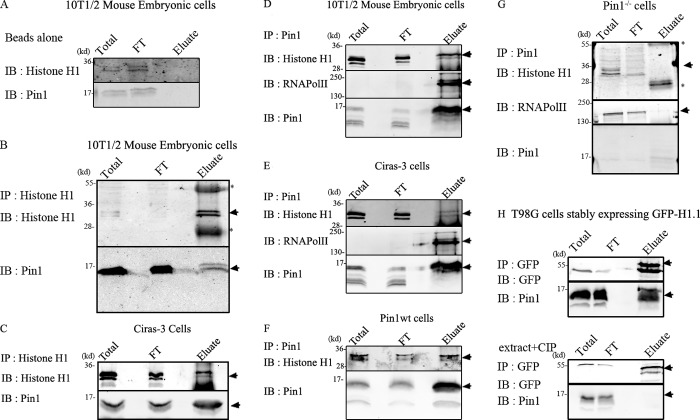
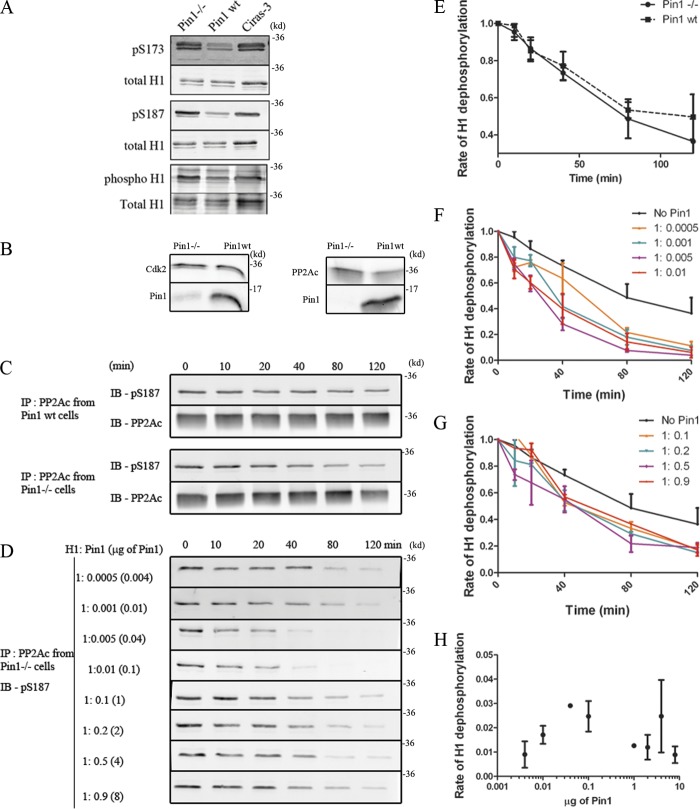
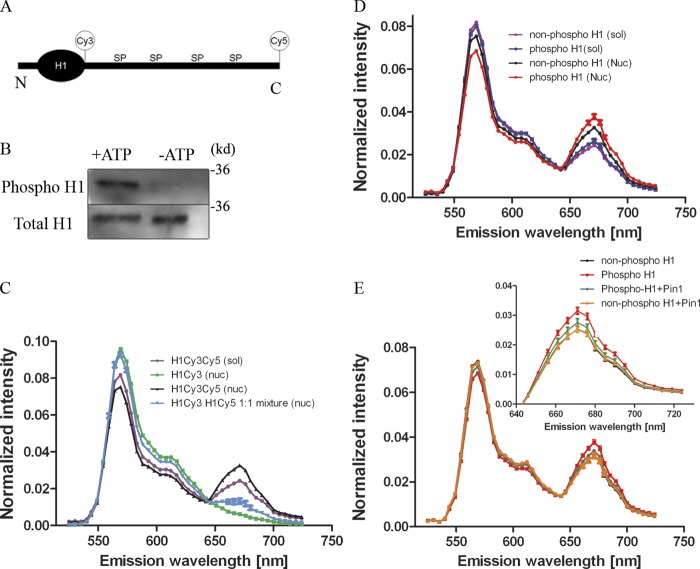
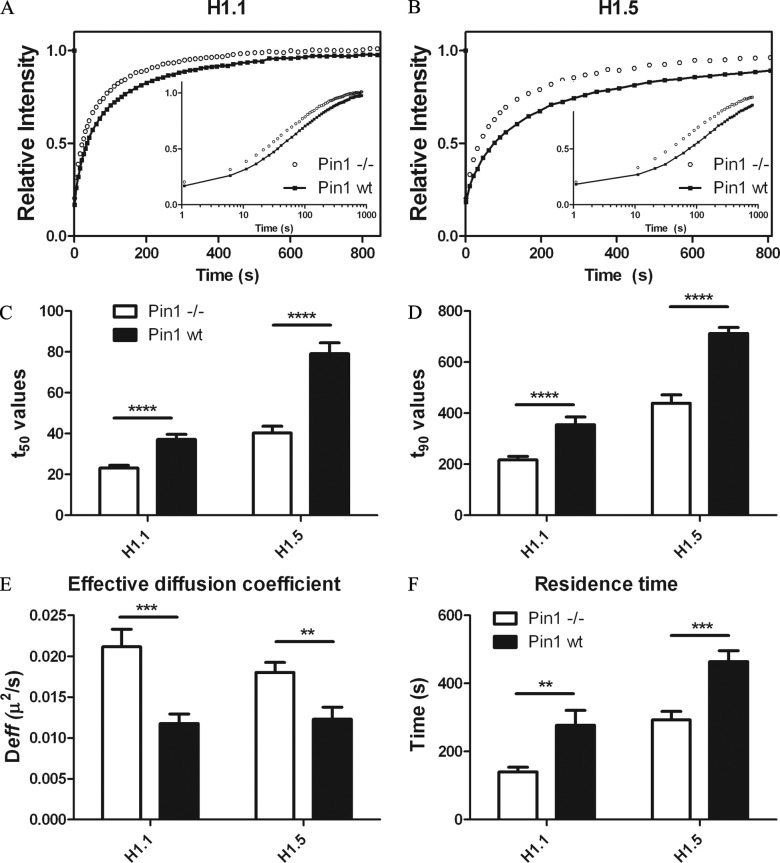
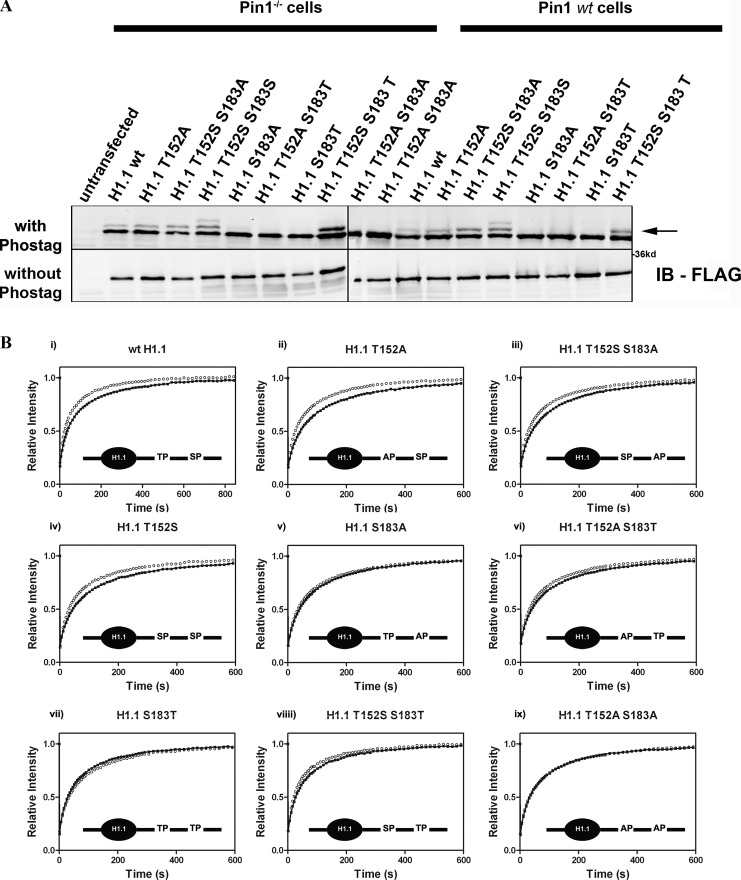
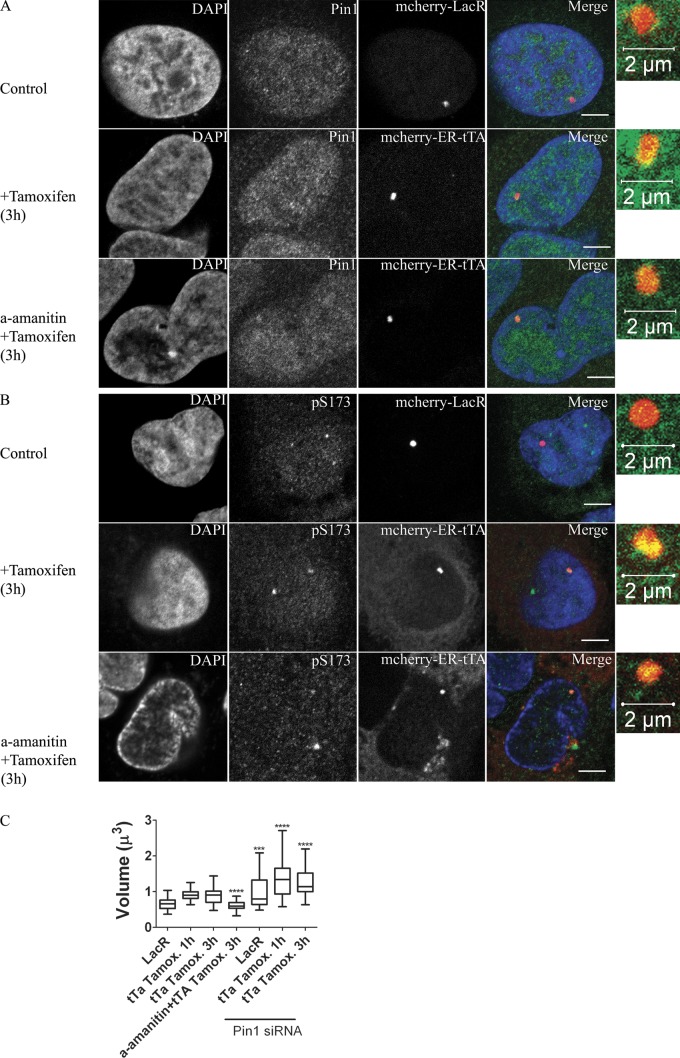
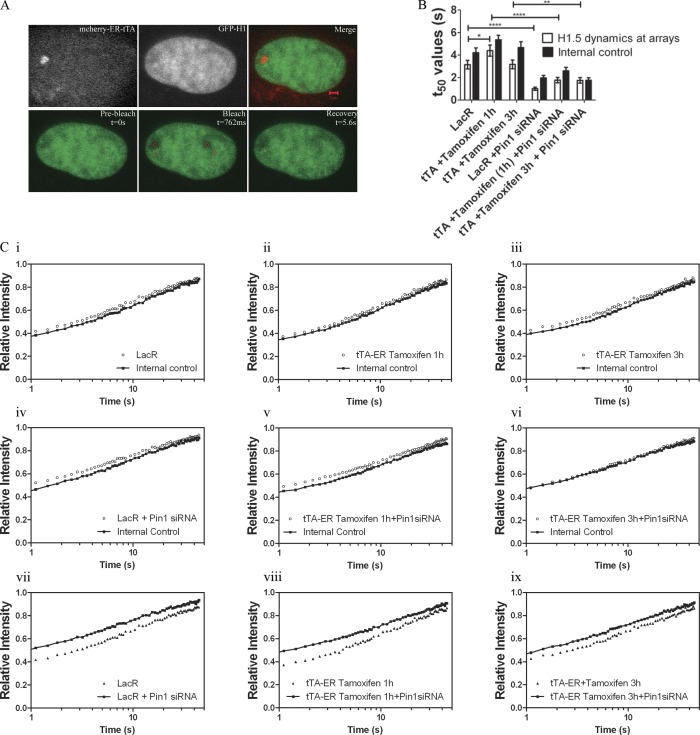
Histone H1 plays a crucial role in stabilizing higher order chromatin structure. Transcriptional activation, DNA replication, and chromosome condensation all require changes in chromatin structure and are correlated with the phosphorylation of histone H1. In this study, we describe a novel interaction between Pin1, a phosphorylation-specific prolyl isomerase, and phosphorylated histone H1. A sub-stoichiometric amount of Pin1 stimulated the dephosphorylation of H1 in vitro and modulated the structure of the C-terminal domain of H1 in a phosphorylation-dependent manner. Depletion of Pin1 destabilized H1 binding to chromatin only when Pin1 binding sites on H1 were present. Pin1 recruitment and localized histone H1 phosphorylation were associated with transcriptional activation independent of RNA polymerase II. We thus identify a novel form of histone H1 regulation through phosphorylation-dependent proline isomerization, which has consequences on overall H1 phosphorylation levels and the stability of H1 binding to chromatin.
Figures
Similar articles
-
Molecular Mechanism of the Pin1-Histone H1 Interaction.Biochemistry. 2019 Feb 12;58(6):788-798. doi: 10.1021/acs.biochem.8b01036. Epub 2018 Dec 18. Biochemistry. 2019. PMID: 30507159 Free PMC article.
-
Peptidylprolyl Isomerase Pin1 Directly Enhances the DNA Binding Functions of Estrogen Receptor α.J Biol Chem. 2015 May 29;290(22):13749-62. doi: 10.1074/jbc.M114.621698. Epub 2015 Apr 12. J Biol Chem. 2015. PMID: 25866209 Free PMC article.
-
Phosphorylation-dependent proline isomerization catalyzed by Pin1 is essential for tumor cell survival and entry into mitosis.Cell Growth Differ. 2000 Jul;11(7):409-16. Cell Growth Differ. 2000. PMID: 10939594
-
Exploring the molecular function of PIN1 by nuclear magnetic resonance.Curr Protein Pept Sci. 2006 Jun;7(3):179-94. doi: 10.2174/138920306777452303. Curr Protein Pept Sci. 2006. PMID: 16787258 Review.
-
Phosphorylation-dependent prolyl isomerization: a novel cell cycle regulatory mechanism.Prog Cell Cycle Res. 2000;4:83-96. doi: 10.1007/978-1-4615-4253-7_8. Prog Cell Cycle Res. 2000. PMID: 10740817 Review.
Cited by
-
Post-translational modifications of the intrinsically disordered terminal domains of histone H1: effects on secondary structure and chromatin dynamics.Chromosoma. 2017 Feb;126(1):83-91. doi: 10.1007/s00412-016-0591-8. Epub 2016 Apr 21. Chromosoma. 2017. PMID: 27098855 Review.
-
Acetylation-modulated communication between the H3 N-terminal tail domain and the intrinsically disordered H1 C-terminal domain.Nucleic Acids Res. 2020 Nov 18;48(20):11510-11520. doi: 10.1093/nar/gkaa949. Nucleic Acids Res. 2020. PMID: 33125082 Free PMC article.
-
Prolyl isomerases in gene transcription.Biochim Biophys Acta. 2015 Oct;1850(10):2017-34. doi: 10.1016/j.bbagen.2014.10.028. Epub 2014 Oct 31. Biochim Biophys Acta. 2015. PMID: 25450176 Free PMC article. Review.
-
Linker histone H1.5 is an underestimated factor in differentiation and carcinogenesis.Environ Epigenet. 2020 Oct 3;6(1):dvaa013. doi: 10.1093/eep/dvaa013. eCollection 2020. Environ Epigenet. 2020. PMID: 33214908 Free PMC article. Review.
-
Proteasome-dependent degradation of histone H1 subtypes is mediated by its C-terminal domain.Protein Sci. 2024 May;33(5):e4970. doi: 10.1002/pro.4970. Protein Sci. 2024. PMID: 38591484 Free PMC article.
References
-
- Albert A., Lavoie S., Vincent M. 1999. A hyperphosphorylated form of RNA polymerase II is the major interphase antigen of the phosphoprotein antibody MPM-2 and interacts with the peptidyl-prolyl isomerase Pin1. J. Cell Sci. 112:2493–2500 - PubMed
-
- Bednar J., Horowitz R.A., Grigoryev S.A., Carruthers L.M., Hansen J.C., Koster A.J., Woodcock C.L. 1998. Nucleosomes, linker DNA, and linker histone form a unique structural motif that directs the higher-order folding and compaction of chromatin. Proc. Natl. Acad. Sci. USA. 95:14173–14178 10.1073/pnas.95.24.14173 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
