

RhoA GTPase controls cytokinesis and programmed necrosis of hematopoietic progenitors
- PMID: 24101377
- PMCID: PMC3804933
- DOI: 10.1084/jem.20122348
RhoA GTPase controls cytokinesis and programmed necrosis of hematopoietic progenitors
Abstract
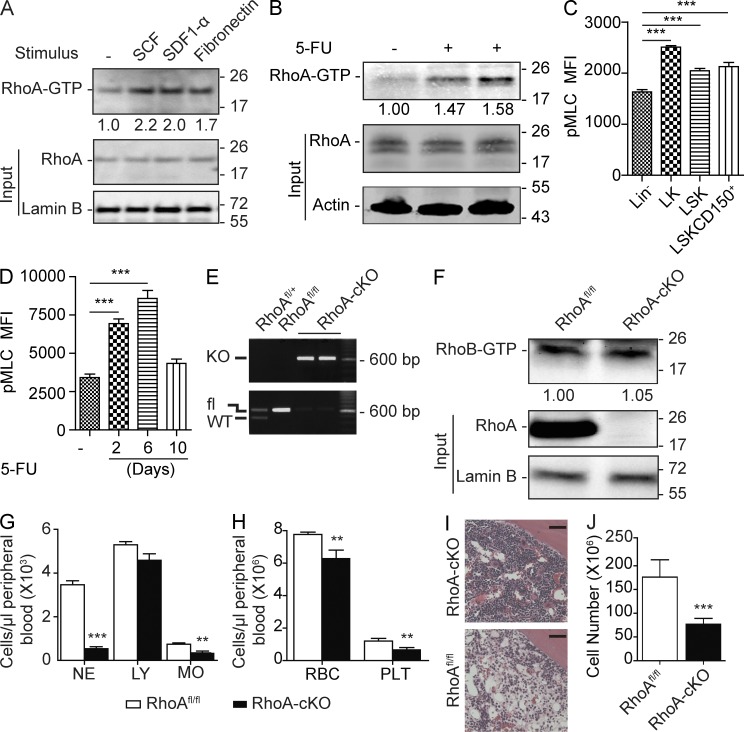
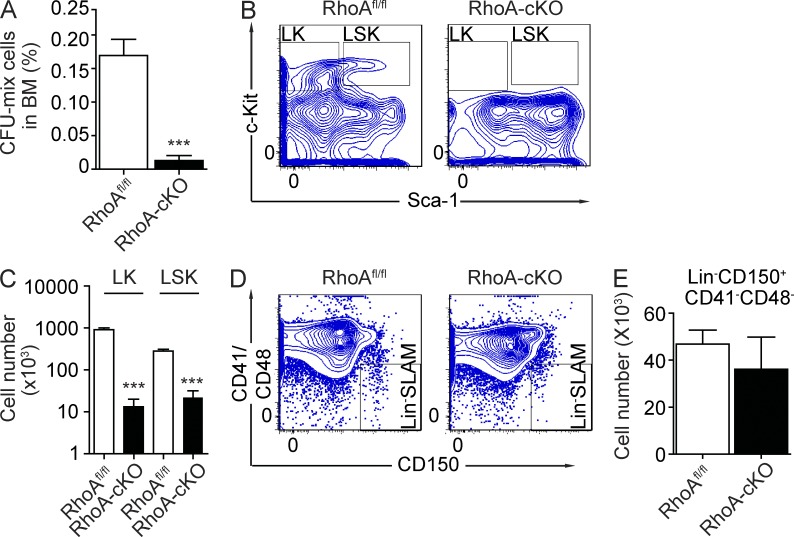
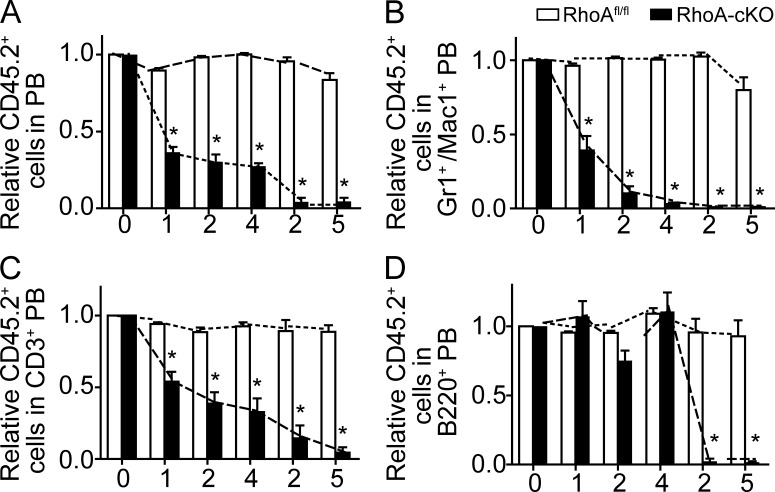
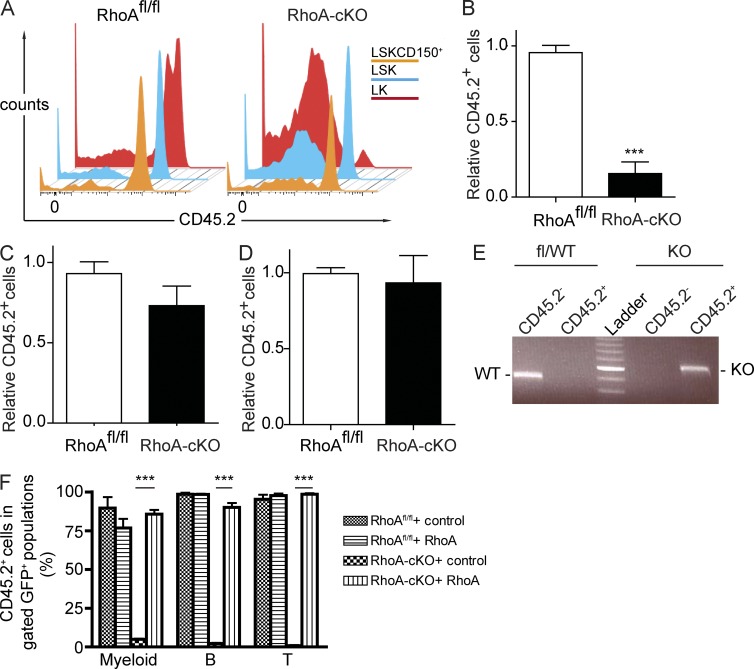
Hematopoietic progenitor cells (HPCs) are central to hematopoiesis as they provide large numbers of lineage-defined blood cells necessary to sustain blood homeostasis. They are one of the most actively cycling somatic cells, and their precise control is critical for hematopoietic homeostasis. The small GTPase RhoA is an intracellular molecular switch that integrates cytokine, chemokine, and adhesion signals to coordinate multiple context-dependent cellular processes. By using a RhoA conditional knockout mouse model, we show that RhoA deficiency causes a multilineage hematopoietic failure that is associated with defective multipotent HPCs. Interestingly, RhoA(-/-) hematopoietic stem cells retained long-term engraftment potential but failed to produce multipotent HPCs and lineage-defined blood cells. This multilineage hematopoietic failure was rescued by reconstituting wild-type RhoA into the RhoA(-/-) Lin(-)Sca-1(+)c-Kit(+) compartment. Mechanistically, RhoA regulates actomyosin signaling, cytokinesis, and programmed necrosis of the HPCs, and loss of RhoA results in a cytokinesis failure of HPCs manifested by an accumulation of multinucleated cells caused by failed abscission of the cleavage furrow after telophase. Concomitantly, the HPCs show a drastically increased death associated with increased TNF-RIP-mediated necrosis. These results show that RhoA is a critical and specific regulator of multipotent HPCs during cytokinesis and thus essential for multilineage hematopoiesis.
Figures
Similar articles
-
In Vivo Rescue Assay of RhoA-Deficient Hematopoietic Stem and Progenitor Cells.Methods Mol Biol. 2018;1821:247-256. doi: 10.1007/978-1-4939-8612-5_17. Methods Mol Biol. 2018. PMID: 30062417 Free PMC article.
-
Cytokinesis failure in RhoA-deficient mouse erythroblasts involves actomyosin and midbody dysregulation and triggers p53 activation.Blood. 2015 Sep 17;126(12):1473-82. doi: 10.1182/blood-2014-12-616169. Epub 2015 Jul 30. Blood. 2015. PMID: 26228485 Free PMC article.
-
Human Cytomegalovirus miR-US25-1 Targets the GTPase RhoA To Inhibit CD34+ Hematopoietic Progenitor Cell Proliferation To Maintain the Latent Viral Genome.mBio. 2021 Apr 6;12(2):e00621-21. doi: 10.1128/mBio.00621-21. mBio. 2021. PMID: 33824207 Free PMC article.
-
Shared mechanisms regulate spatiotemporal RhoA-dependent actomyosin contractility during adhesion and cell division.Small GTPases. 2020 Mar;11(2):113-121. doi: 10.1080/21541248.2017.1366966. Epub 2017 Dec 31. Small GTPases. 2020. PMID: 29291271 Free PMC article. Review.
-
RHO subfamily of small GTPases in the development and function of hematopoietic cells.J Cell Physiol. 2025 Jan;240(1):e31469. doi: 10.1002/jcp.31469. Epub 2024 Oct 21. J Cell Physiol. 2025. PMID: 39434451 Review.
Cited by
-
RHOA Therapeutic Targeting in Hematological Cancers.Cells. 2023 Jan 28;12(3):433. doi: 10.3390/cells12030433. Cells. 2023. PMID: 36766776 Free PMC article. Review.
-
Inhibition of RHOA activity preserves the survival and hemostasis function of long-term cold-stored platelets.Blood. 2024 Oct 17;144(16):1732-1746. doi: 10.1182/blood.2023021453. Blood. 2024. PMID: 39088777
-
α-Catenin force-sensitive binding and sequestration of LZTS2 leads to cytokinesis failure.J Cell Biol. 2025 Mar 3;224(3):e202308124. doi: 10.1083/jcb.202308124. Epub 2025 Jan 9. J Cell Biol. 2025. PMID: 39786338
-
PRDM16 isoforms differentially regulate normal and leukemic hematopoiesis and inflammatory gene signature.J Clin Invest. 2018 Aug 1;128(8):3250-3264. doi: 10.1172/JCI99862. Epub 2018 Jul 23. J Clin Invest. 2018. PMID: 29878897 Free PMC article.
-
Deconstructing the Complexity of TGFβ Signaling in Hematopoietic Stem Cells: Quiescence and Beyond.Curr Stem Cell Rep. 2016 Dec;2(4):388-397. doi: 10.1007/s40778-016-0069-x. Epub 2016 Oct 29. Curr Stem Cell Rep. 2016. PMID: 28529843 Free PMC article.
References
-
- Boggs D.R., Boggs S.S. 1976. Editorial: The pathogenesis of aplastic anemia: a defective pluripotent hematopoietic stem cell with inappropriate balance of differentiation and self-replication. Blood. 48:71–76 - PubMed
-
- Broudy V.C. 1997. Stem cell factor and hematopoiesis. Blood. 90:1345–1364 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
