

Cellular organization of cortical barrel columns is whisker-specific
- PMID: 24101458
- PMCID: PMC3839692
- DOI: 10.1073/pnas.1312691110
Cellular organization of cortical barrel columns is whisker-specific
Abstract
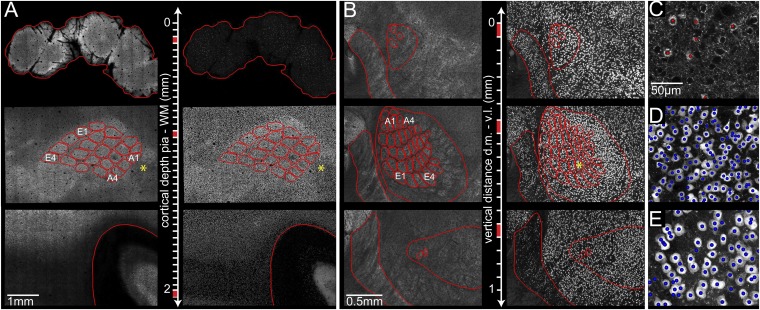
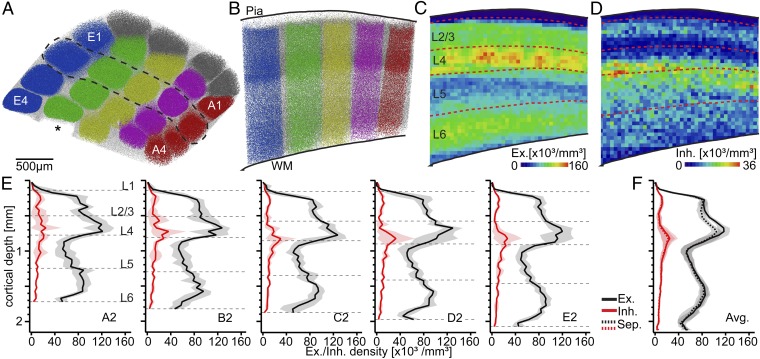
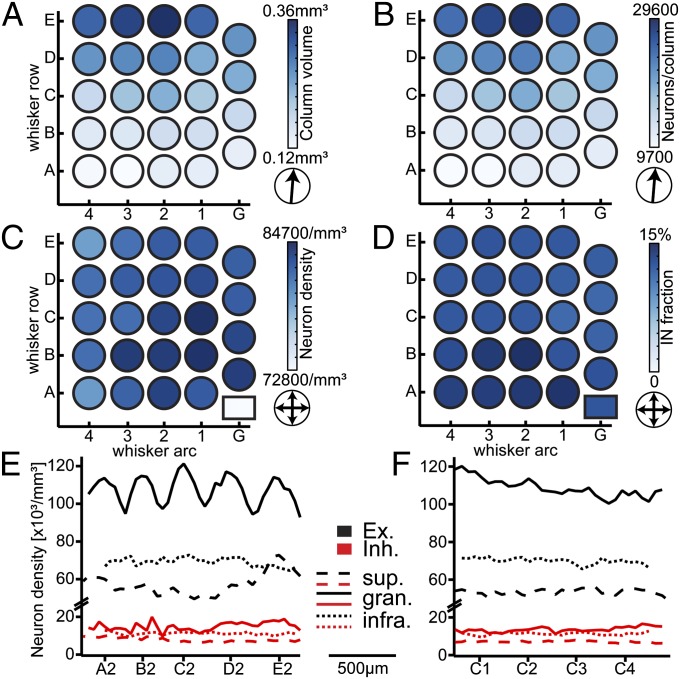
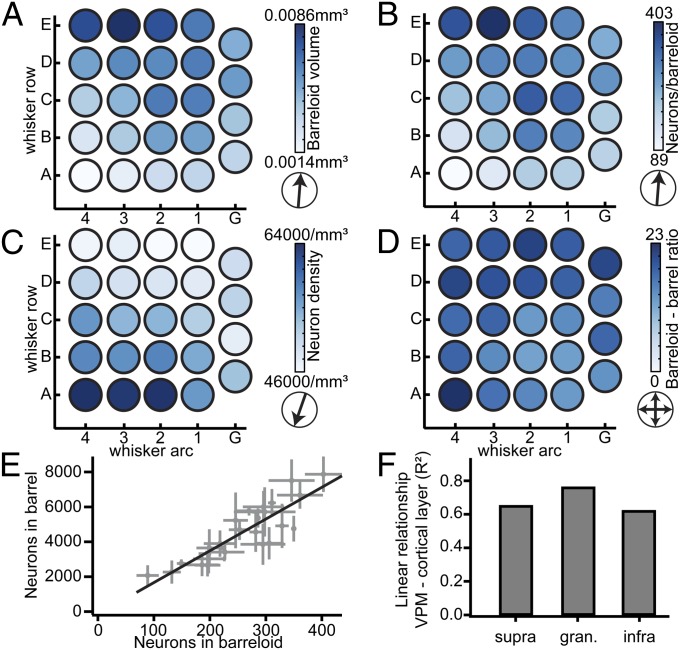
The cellular organization of the cortex is of fundamental importance for elucidating the structural principles that underlie its functions. It has been suggested that reconstructing the structure and synaptic wiring of the elementary functional building block of mammalian cortices, the cortical column, might suffice to reverse engineer and simulate the functions of entire cortices. In the vibrissal area of rodent somatosensory cortex, whisker-related "barrel" columns have been referred to as potential cytoarchitectonic equivalents of functional cortical columns. Here, we investigated the structural stereotypy of cortical barrel columns by measuring the 3D neuronal composition of the entire vibrissal area in rat somatosensory cortex and thalamus. We found that the number of neurons per cortical barrel column and thalamic "barreloid" varied substantially within individual animals, increasing by ∼2.5-fold from dorsal to ventral whiskers. As a result, the ratio between whisker-specific thalamic and cortical neurons was remarkably constant. Thus, we hypothesize that the cellular architecture of sensory cortices reflects the degree of similarity in sensory input and not columnar and/or cortical uniformity principles.
Keywords: GAD67; NeuN; VPM; barrel cortex; soma counts.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Comment in
-
Cerebral cortex: Sizing up the columns.Nat Rev Neurosci. 2013 Dec;14(12):817. doi: 10.1038/nrn3631. Epub 2013 Oct 30. Nat Rev Neurosci. 2013. PMID: 24169936 No abstract available.
Similar articles
-
Beyond Columnar Organization: Cell Type- and Target Layer-Specific Principles of Horizontal Axon Projection Patterns in Rat Vibrissal Cortex.Cereb Cortex. 2015 Nov;25(11):4450-68. doi: 10.1093/cercor/bhv053. Epub 2015 Apr 1. Cereb Cortex. 2015. PMID: 25838038 Free PMC article.
-
Functional topography of corticothalamic feedback enhances thalamic spatial response tuning in the somatosensory whisker/barrel system.Neuron. 2004 Feb 19;41(4):639-51. doi: 10.1016/s0896-6273(04)00046-7. Neuron. 2004. PMID: 14980211
-
Spatiotemporal dynamics of sensory responses in layer 2/3 of rat barrel cortex measured in vivo by voltage-sensitive dye imaging combined with whole-cell voltage recordings and neuron reconstructions.J Neurosci. 2003 Feb 15;23(4):1298-309. doi: 10.1523/JNEUROSCI.23-04-01298.2003. J Neurosci. 2003. PMID: 12598618 Free PMC article.
-
Excitatory signal flow and connectivity in a cortical column: focus on barrel cortex.Brain Struct Funct. 2007 Jul;212(1):3-17. doi: 10.1007/s00429-007-0144-2. Epub 2007 Jun 1. Brain Struct Funct. 2007. PMID: 17717695 Review.
-
Barrel cortex function.Prog Neurobiol. 2013 Apr;103:3-27. doi: 10.1016/j.pneurobio.2012.11.002. Epub 2012 Nov 27. Prog Neurobiol. 2013. PMID: 23195880 Review.
Cited by
-
Neural Substrate Expansion for the Restoration of Brain Function.Front Syst Neurosci. 2016 Jan 25;10:1. doi: 10.3389/fnsys.2016.00001. eCollection 2016. Front Syst Neurosci. 2016. PMID: 26834579 Free PMC article. Review.
-
Cerebral cortex: Sizing up the columns.Nat Rev Neurosci. 2013 Dec;14(12):817. doi: 10.1038/nrn3631. Epub 2013 Oct 30. Nat Rev Neurosci. 2013. PMID: 24169936 No abstract available.
-
Three-dimensional scanless holographic optogenetics with temporal focusing (3D-SHOT).Nat Commun. 2017 Oct 31;8(1):1228. doi: 10.1038/s41467-017-01031-3. Nat Commun. 2017. PMID: 29089483 Free PMC article.
-
Sparse connectivity enables efficient information processing in cortex-like artificial neural networks.Front Neural Circuits. 2025 Mar 13;19:1528309. doi: 10.3389/fncir.2025.1528309. eCollection 2025. Front Neural Circuits. 2025. PMID: 40182663 Free PMC article.
-
Invariant visual object recognition and shape processing in rats.Behav Brain Res. 2015 May 15;285:10-33. doi: 10.1016/j.bbr.2014.12.053. Epub 2015 Jan 2. Behav Brain Res. 2015. PMID: 25561421 Free PMC article. Review.
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
