

Allosteric regulation of SERCA by phosphorylation-mediated conformational shift of phospholamban
- PMID: 24101520
- PMCID: PMC3808617
- DOI: 10.1073/pnas.1303006110
Allosteric regulation of SERCA by phosphorylation-mediated conformational shift of phospholamban
Abstract
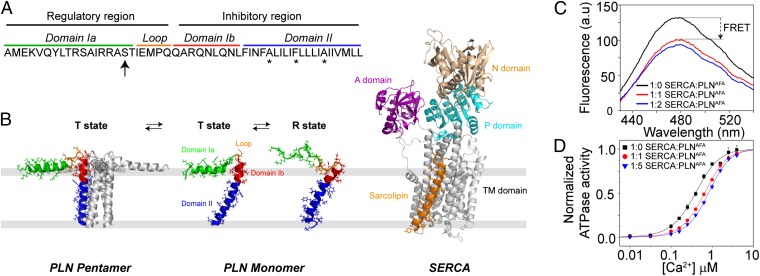
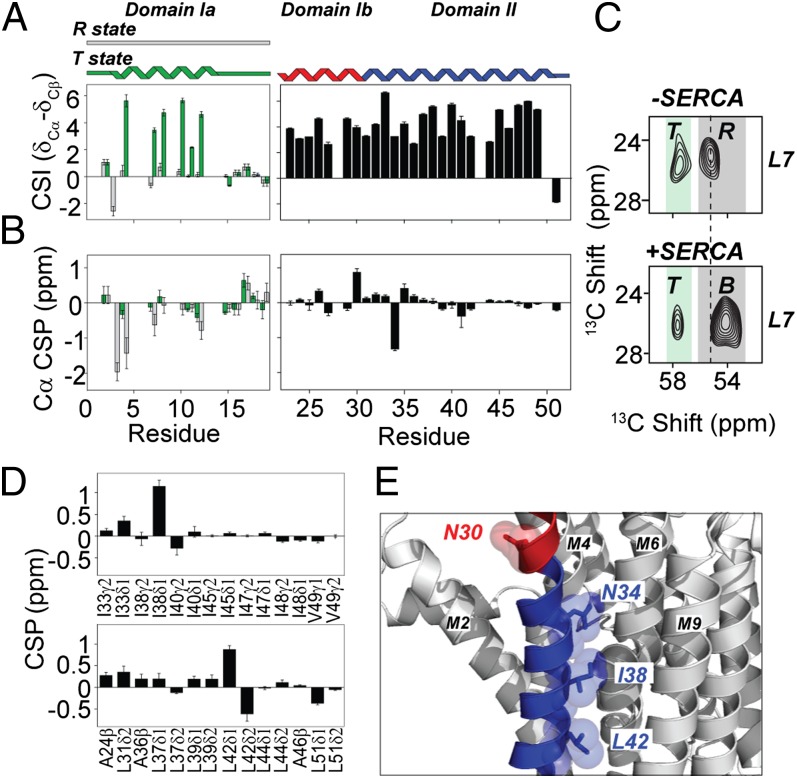
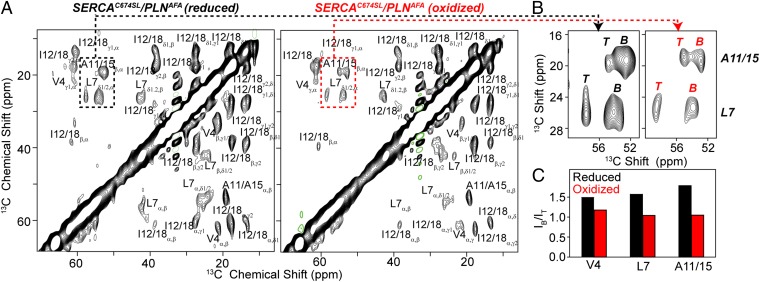
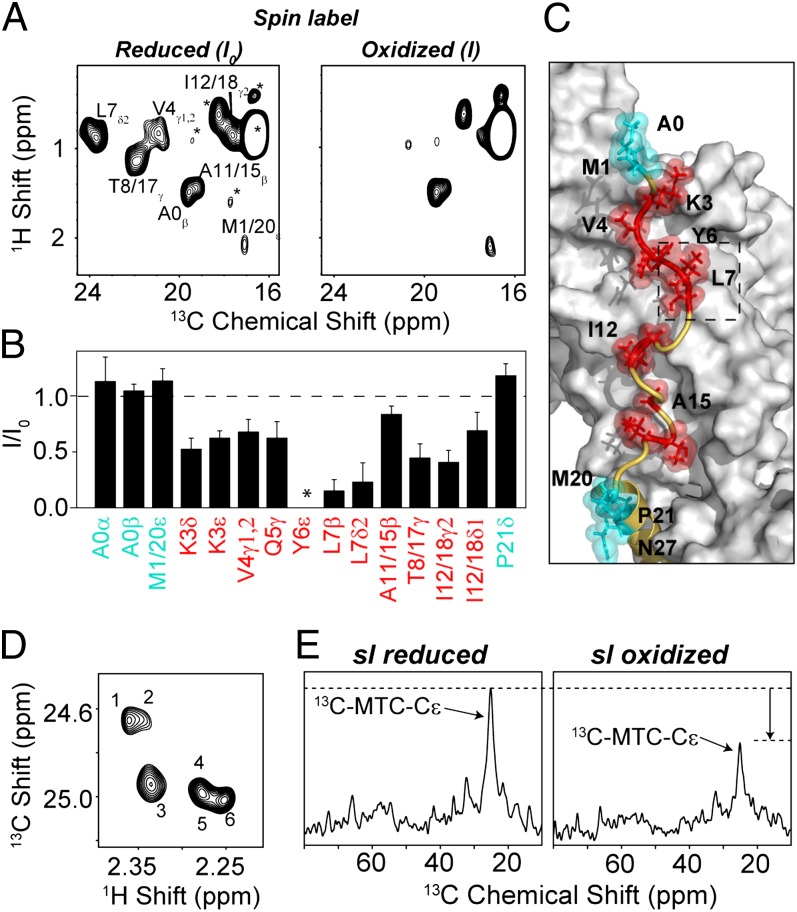
The membrane protein complex between the sarcoplasmic reticulum Ca(2+)-ATPase (SERCA) and phospholamban (PLN) controls Ca(2+) transport in cardiomyocytes, thereby modulating cardiac contractility. β-Adrenergic-stimulated phosphorylation of PLN at Ser-16 enhances SERCA activity via an unknown mechanism. Using solid-state nuclear magnetic resonance spectroscopy, we mapped the physical interactions between SERCA and both unphosphorylated and phosphorylated PLN in membrane bilayers. We found that the allosteric regulation of SERCA depends on the conformational equilibrium of PLN, whose cytoplasmic regulatory domain interconverts between three different states: a ground T state (helical and membrane associated), an excited R state (unfolded and membrane detached), and a B state (extended and enzyme-bound), which is noninhibitory. Phosphorylation at Ser-16 of PLN shifts the populations toward the B state, increasing SERCA activity. We conclude that PLN's conformational equilibrium is central to maintain SERCA's apparent Ca(2+) affinity within a physiological window. This model represents a paradigm shift in our understanding of SERCA regulation by posttranslational phosphorylation and suggests strategies for designing innovative therapeutic approaches to enhance cardiac muscle contractility.
Keywords: magic angle spinning; paramagnetic relaxation enhancement; protein-protein interactions; solid-state NMR.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
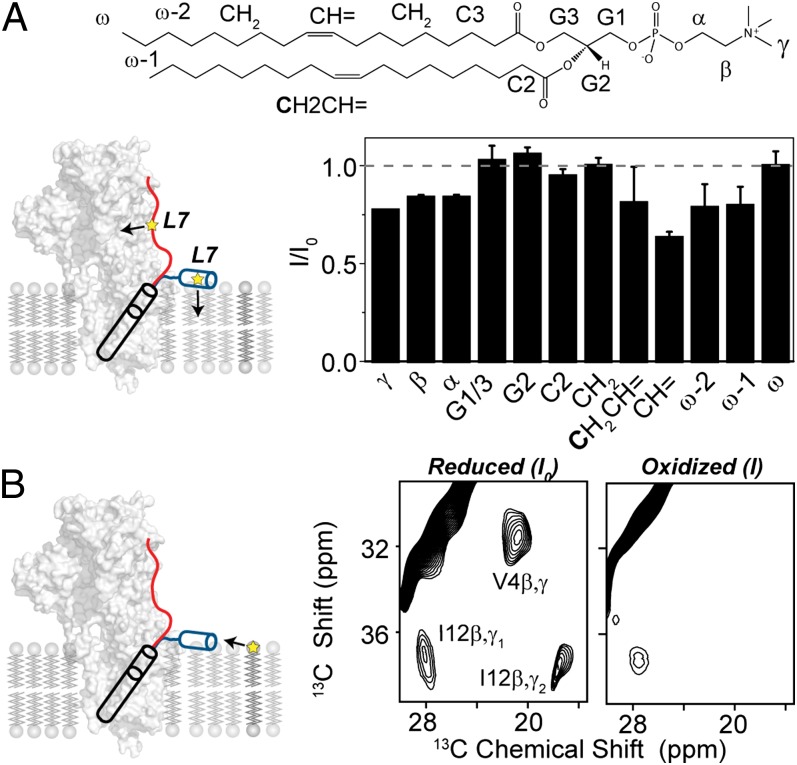
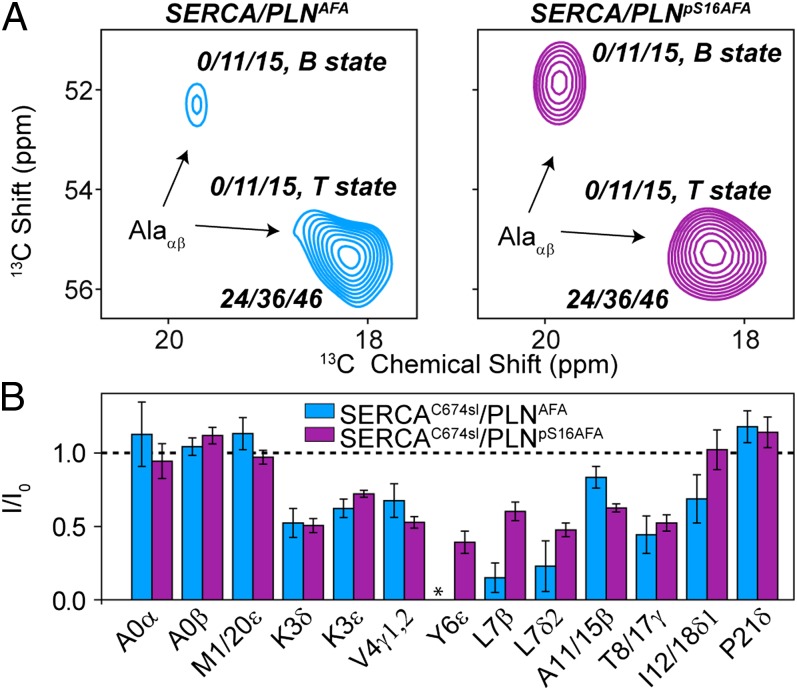
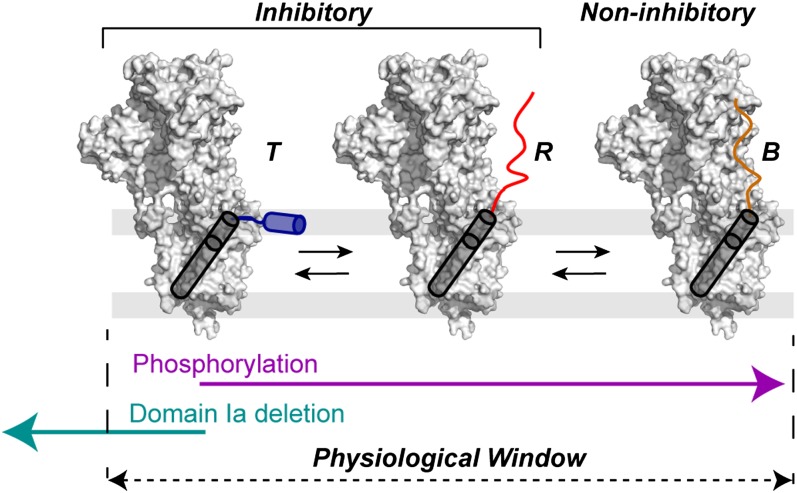
References
-
- Bers DM. Cardiac excitation-contraction coupling. Nature. 2002;415(6868):198–205. - PubMed
-
- Brini M, Carafoli E. Calcium pumps in health and disease. Physiol Rev. 2009;89(4):1341–1378. - PubMed
-
- MacLennan DH, Kranias EG. Phospholamban: A crucial regulator of cardiac contractility. Nat Rev Mol Cell Biol. 2003;4(7):566–577. - PubMed
-
- Toyoshima C, Nakasako M, Nomura H, Ogawa H. Crystal structure of the calcium pump of sarcoplasmic reticulum at 2.6 A resolution. Nature. 2000;405(6787):647–655. - PubMed
-
- Møller JV, Olesen C, Winther AM, Nissen P. The sarcoplasmic Ca2+-ATPase: design of a perfect chemi-osmotic pump. Q Rev Biophys. 2010;43(4):501–566. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
