

A method for the three-dimensional reconstruction of Neurobiotin™-filled neurons and the location of their synaptic inputs
- PMID: 24101895
- PMCID: PMC3787200
- DOI: 10.3389/fncir.2013.00153
A method for the three-dimensional reconstruction of Neurobiotin™-filled neurons and the location of their synaptic inputs
Abstract
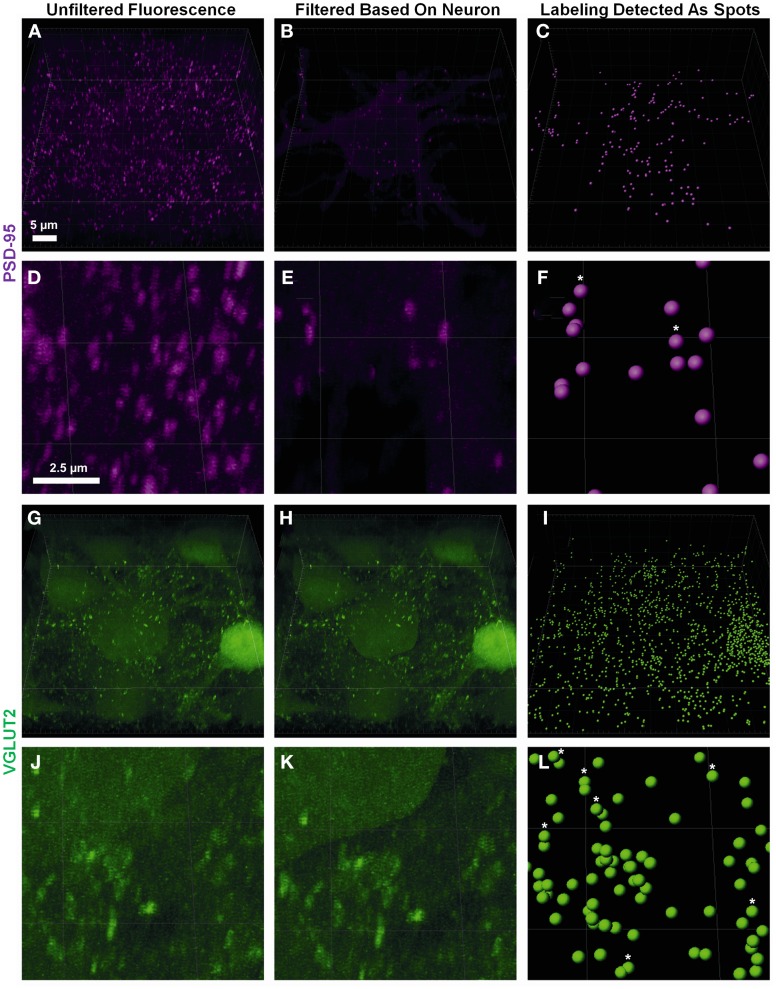
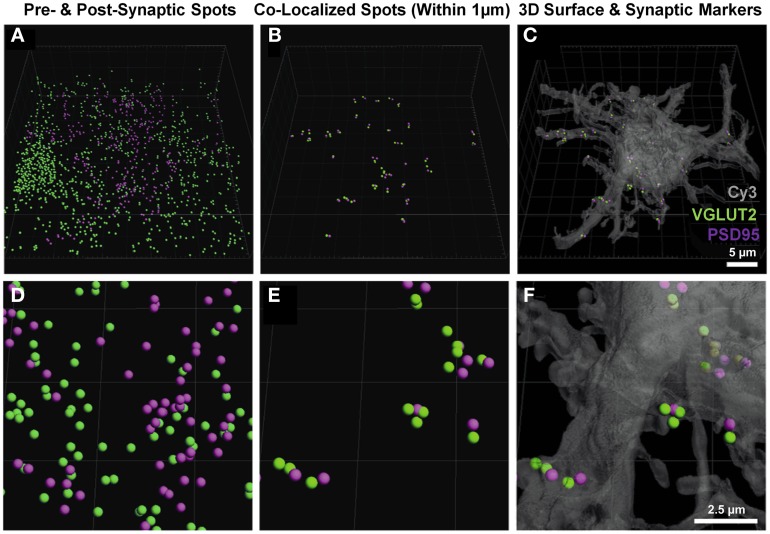
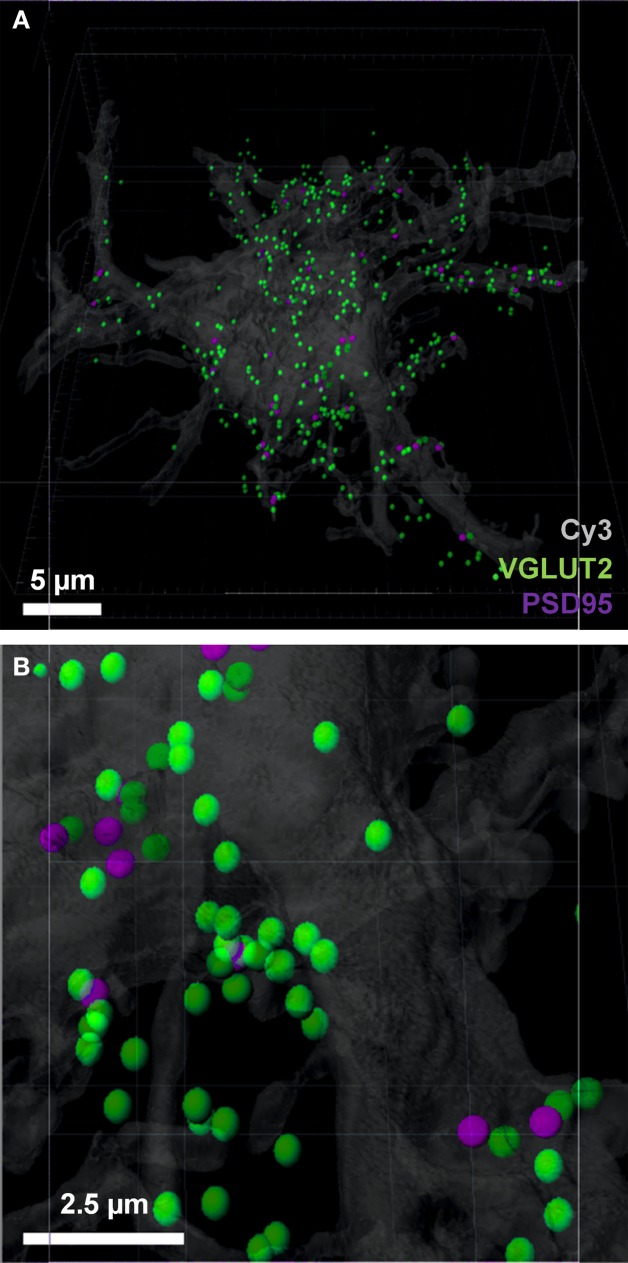
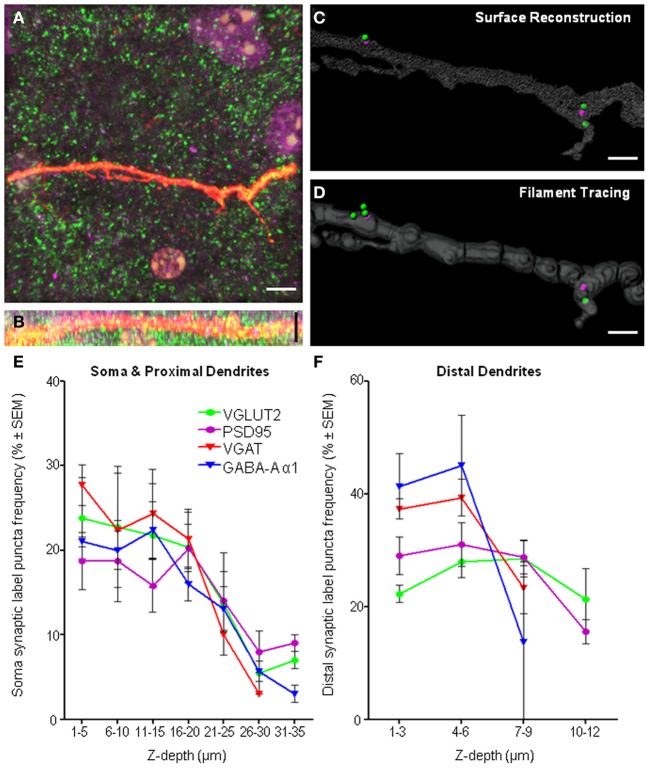
Here, we describe a robust method for mapping the number and type of neuro-chemically distinct synaptic inputs that a single reconstructed neuron receives. We have used individual hypoglossal motor neurons filled with Neurobiotin by semi-loose seal electroporation in thick brainstem slices. These filled motor neurons were then processed for excitatory and inhibitory synaptic inputs, using immunohistochemical-labeling procedures. For excitatory synapses, we used anti-VGLUT2 to locate glutamatergic pre-synaptic terminals and anti-PSD-95 to locate post-synaptic specializations on and within the surface of these filled motor neurons. For inhibitory synapses, we used anti-VGAT to locate GABAergic pre-synaptic terminals and anti-GABA-A receptor subunit α1 to locate the post-synaptic domain. The Neurobiotin-filled and immuno-labeled motor neuron was then processed for optical sectioning using confocal microscopy. The morphology of the motor neuron including its dendritic tree and the distribution of excitatory and inhibitory synapses were then determined by three-dimensional reconstruction using IMARIS software (Bitplane). Using surface rendering, fluorescence thresholding, and masking of unwanted immuno-labeling, tools found in IMARIS, we were able to obtain an accurate 3D structure of an individual neuron including the number and location of its glutamatergic and GABAergic synaptic inputs. The power of this method allows for a rapid morphological confirmation of the post-synaptic responses recorded by patch-clamp prior to Neurobiotin filling. Finally, we show that this method can be adapted to super-resolution microscopy techniques, which will enhance its applicability to the study of neural circuits at the level of synapses.
Keywords: 3D-reconstruction; brainstem; dendrite; motor neuron; synapse.
Figures




Similar articles
-
Alterations in hypoglossal motor neurons due to GAD67 and VGAT deficiency in mice.Exp Neurol. 2017 Mar;289:117-127. doi: 10.1016/j.expneurol.2016.12.004. Epub 2016 Dec 10. Exp Neurol. 2017. PMID: 27956032
-
Glycinergic Neurotransmission: A Potent Regulator of Embryonic Motor Neuron Dendritic Morphology and Synaptic Plasticity.J Neurosci. 2016 Jan 6;36(1):80-7. doi: 10.1523/JNEUROSCI.1576-15.2016. J Neurosci. 2016. PMID: 26740651 Free PMC article.
-
Localization of presynaptic inputs on dendrites of individually labeled neurons in three dimensional space using a center distance algorithm.J Neurosci Methods. 2011 Sep 15;200(2):129-43. doi: 10.1016/j.jneumeth.2011.06.021. Epub 2011 Jun 25. J Neurosci Methods. 2011. PMID: 21736898
-
Synaptic integration in dendritic trees.J Neurobiol. 2005 Jul;64(1):75-90. doi: 10.1002/neu.20144. J Neurobiol. 2005. PMID: 15884003 Review.
-
Synaptic integration in motoneurons with hyper-excitable dendrites.Can J Physiol Pharmacol. 2004 Aug-Sep;82(8-9):549-55. doi: 10.1139/y04-046. Can J Physiol Pharmacol. 2004. PMID: 15523512 Review.
Cited by
-
Hyperglycemia- and neuropathy-induced changes in mitochondria within sensory nerves.Ann Clin Transl Neurol. 2014 Oct;1(10):799-812. doi: 10.1002/acn3.119. Epub 2014 Oct 1. Ann Clin Transl Neurol. 2014. PMID: 25493271 Free PMC article.
-
Familiarity with a vocal category biases the compartmental expression of Arc/Arg3.1 in core auditory cortex.Learn Mem. 2017 Nov 15;24(12):612-621. doi: 10.1101/lm.046086.117. Print 2017 Dec. Learn Mem. 2017. PMID: 29142056 Free PMC article.
-
From initial formation to developmental refinement: GABAergic inputs shape neuronal subnetworks in the primary somatosensory cortex.iScience. 2025 Feb 26;28(3):112104. doi: 10.1016/j.isci.2025.112104. eCollection 2025 Mar 21. iScience. 2025. PMID: 40129704 Free PMC article.
-
Fluoxetine-induced recovery of serotonin and norepinephrine projections in a mouse model of post-stroke depression.Transl Psychiatry. 2020 Sep 30;10(1):334. doi: 10.1038/s41398-020-01008-9. Transl Psychiatry. 2020. PMID: 32999279 Free PMC article.
-
Analysis of Motor Neurons Differentiated from Human Induced Pluripotent Stem Cells for the Use in Cell-Based Botulinum Neurotoxin Activity Assays.Toxins (Basel). 2020 Apr 25;12(5):276. doi: 10.3390/toxins12050276. Toxins (Basel). 2020. PMID: 32344847 Free PMC article.
References
-
- Bae Y. C., Nakamura T., Ihn H. J., Choi M. H., Yoshida A., Moritani M., et al. (1999). Distribution pattern of inhibitory and excitatory synapses in the dendritic tree of single masseter alpha-motoneurons in the cat. J. Comp. Neurol. 414, 454–468 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources