

Molecular logic of neocortical projection neuron specification, development and diversity
- PMID: 24105342
- PMCID: PMC3876965
- DOI: 10.1038/nrn3586
Molecular logic of neocortical projection neuron specification, development and diversity
Abstract
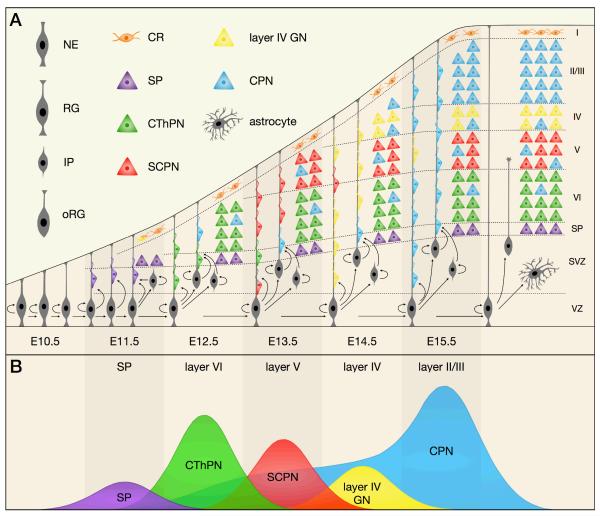
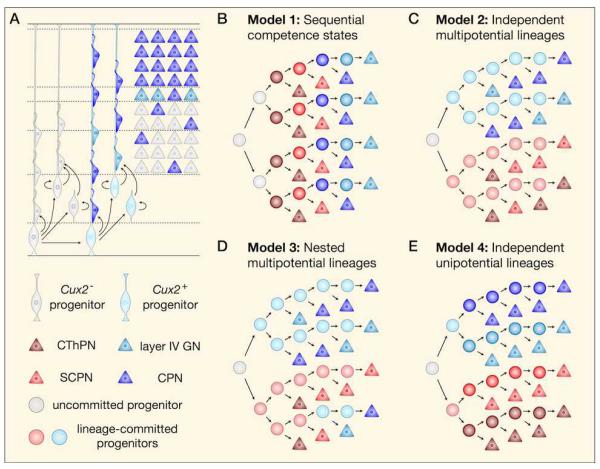
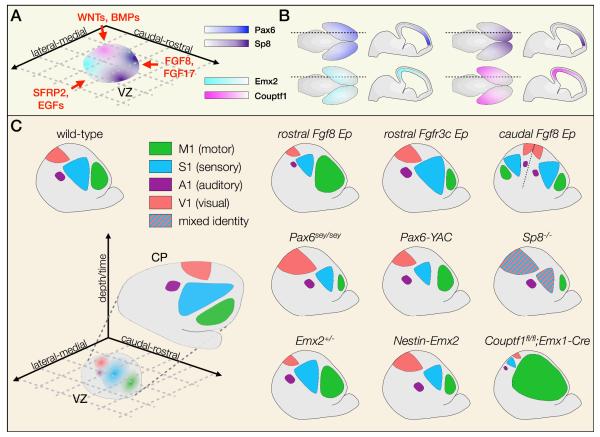
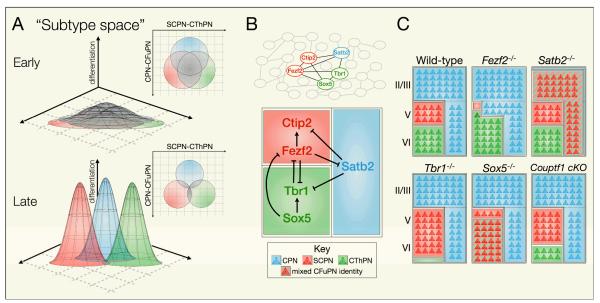
The sophisticated circuitry of the neocortex is assembled from a diverse repertoire of neuronal subtypes generated during development under precise molecular regulation. In recent years, several key controls over the specification and differentiation of neocortical projection neurons have been identified. This work provides substantial insight into the 'molecular logic' underlying cortical development and increasingly supports a model in which individual progenitor-stage and postmitotic regulators are embedded within highly interconnected networks that gate sequential developmental decisions. Here, we provide an integrative account of the molecular controls that direct the progressive development and delineation of subtype and area identity of neocortical projection neurons.
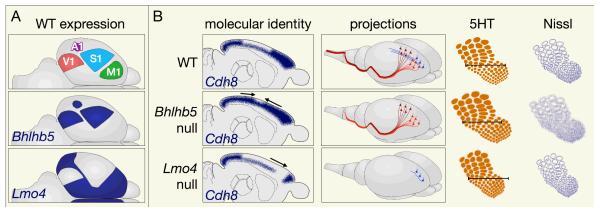
Figures
References
-
- Parnavelas JG. The origin and migration of cortical neurones: new vistas. Trends Neurosci. 2000;23:126–131. - PubMed
-
- Wichterle H, Turnbull DH, Nery S, Fishell G, Alvarez-Buylla A. In utero fate mapping reveals distinct migratory pathways and fates of neurons born in the mammalian basal forebrain. Development. 2001;128:3759–3771. - PubMed
-
- Cobos I, Puelles L, Martinez S. The avian telencephalic subpallium originates inhibitory neurons that invade tangentially the pallium (dorsal ventricular ridge and cortical areas) Dev Biol. 2001;239:30–45. - PubMed
-
- Wonders CP, Anderson SA. The origin and specification of cortical interneurons. Nat Rev Neurosci. 2006;7:687–696. - PubMed
Publication types
MeSH terms
Grants and funding
- NS064730/NS/NINDS NIH HHS/United States
- R01 NS045523/NS/NINDS NIH HHS/United States
- F31 NS080343/NS/NINDS NIH HHS/United States
- R01 NS075672/NS/NINDS NIH HHS/United States
- NS045523/NS/NINDS NIH HHS/United States
- R01 NS049553/NS/NINDS NIH HHS/United States
- NS041590/NS/NINDS NIH HHS/United States
- NS049553/NS/NINDS NIH HHS/United States
- NS075672/NS/NINDS NIH HHS/United States
- NS080343/NS/NINDS NIH HHS/United States
- R37 NS041590/NS/NINDS NIH HHS/United States
- F31 NS064730/NS/NINDS NIH HHS/United States
- T32 GM007753/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
