

A large-scale neural network model of the influence of neuromodulatory levels on working memory and behavior
- PMID: 24106474
- PMCID: PMC3789270
- DOI: 10.3389/fncom.2013.00133
A large-scale neural network model of the influence of neuromodulatory levels on working memory and behavior
Abstract
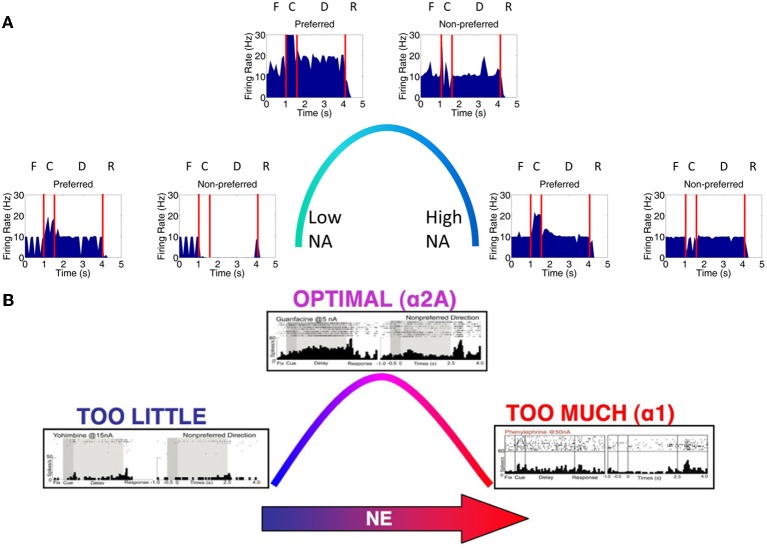
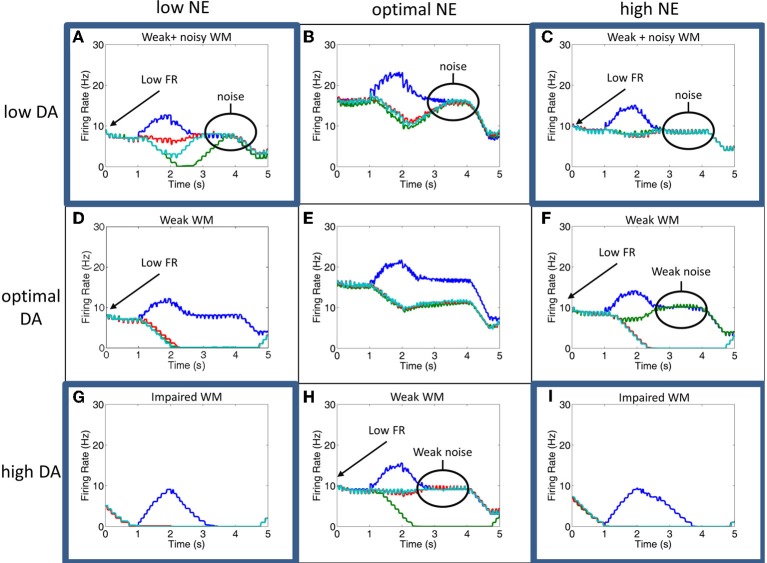
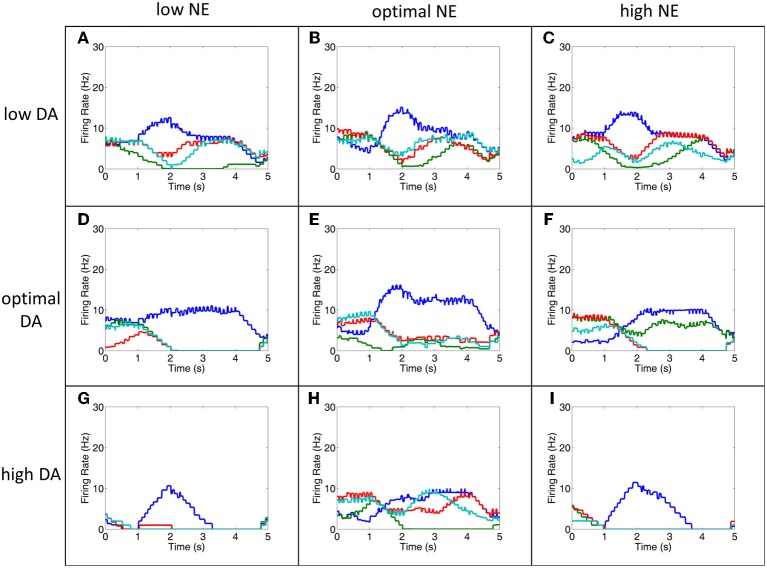
The dorsolateral prefrontal cortex (dlPFC), which is regarded as the primary site for visuospatial working memory in the brain, is significantly modulated by dopamine (DA) and norepinephrine (NE). DA and NE originate in the ventral tegmental area (VTA) and locus coeruleus (LC), respectively, and have been shown to have an "inverted-U" dose-response profile in dlPFC, where the level of arousal and decision-making performance is a function of DA and NE concentrations. Moreover, there appears to be a sweet spot, in terms of the level of DA and NE activation, which allows for optimal working memory and behavioral performance. When either DA or NE is too high, input to the PFC is essentially blocked. When either DA or NE is too low, PFC network dynamics become noisy and activity levels diminish. Mechanisms for how this is occurring have been suggested, however, they have not been tested in a large-scale model with neurobiologically plausible network dynamics. Also, DA and NE levels have not been simultaneously manipulated experimentally, which is not realistic in vivo due to strong bi-directional connections between the VTA and LC. To address these issues, we built a spiking neural network model that includes D1, α2A, and α1 receptors. The model was able to match the inverted-U profiles that have been shown experimentally for differing levels of DA and NE. Furthermore, we were able to make predictions about what working memory and behavioral deficits may occur during simultaneous manipulation of DA and NE outside of their optimal levels. Specifically, when DA levels were low and NE levels were high, cues could not be held in working memory due to increased noise. On the other hand, when DA levels were high and NE levels were low, incorrect decisions were made due to weak overall network activity. We also show that lateral inhibition in working memory may play a more important role in increasing signal-to-noise ratio than increasing recurrent excitatory input.
Keywords: dopamine; neuromodulation; noradrenaline; spiking neural networks; working memory.
Figures
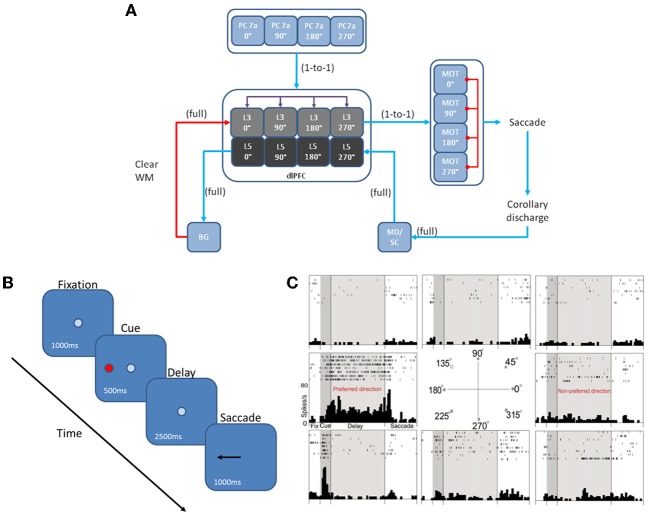
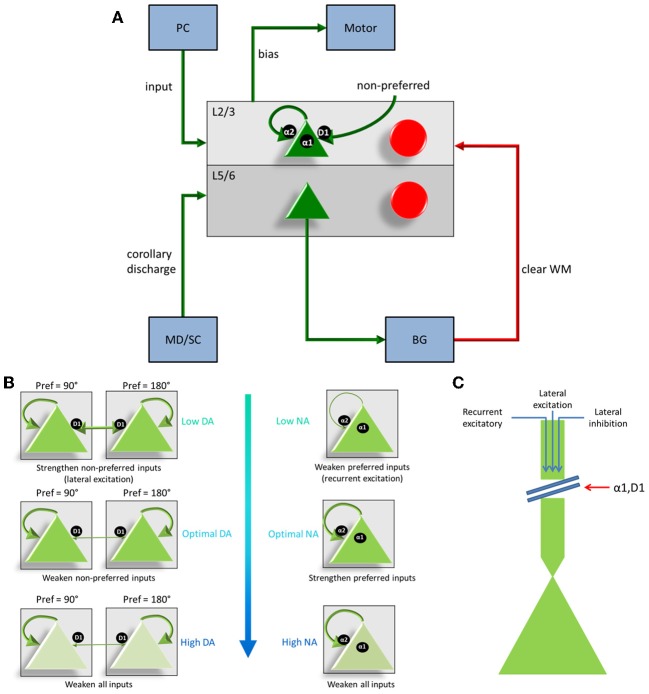
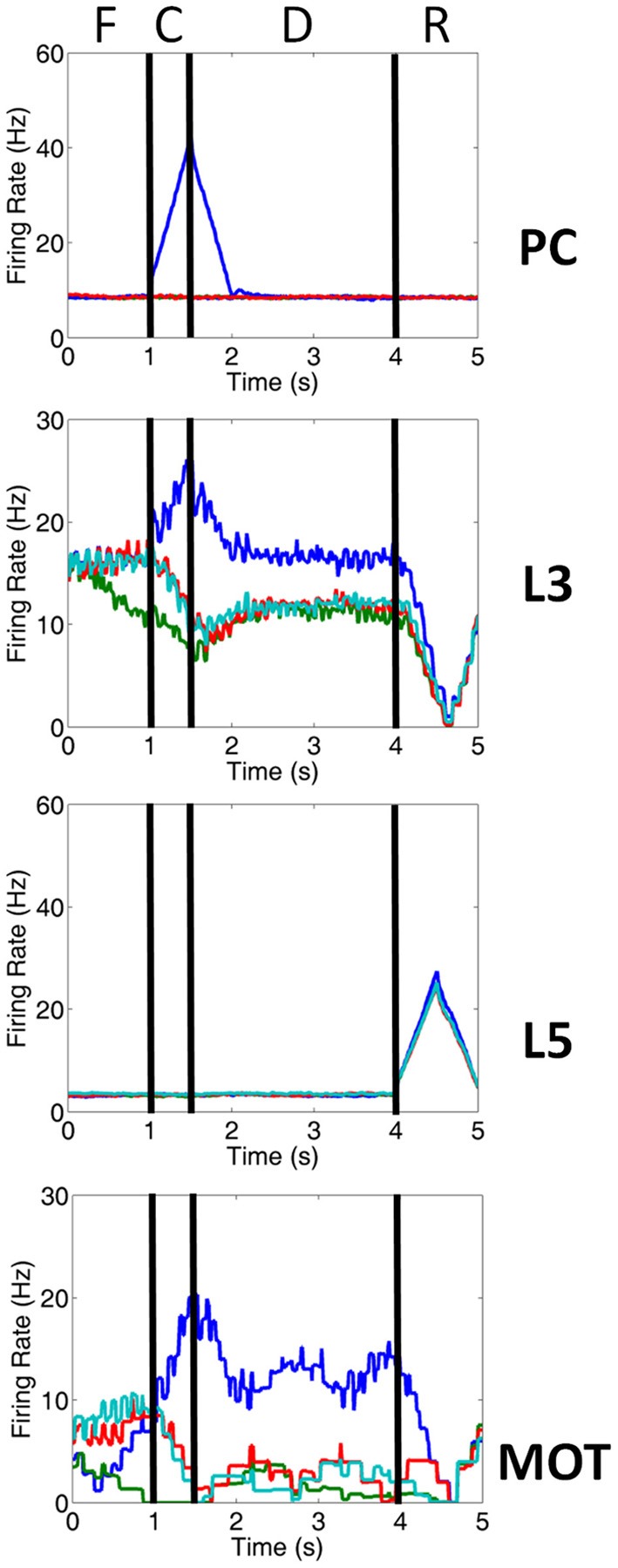
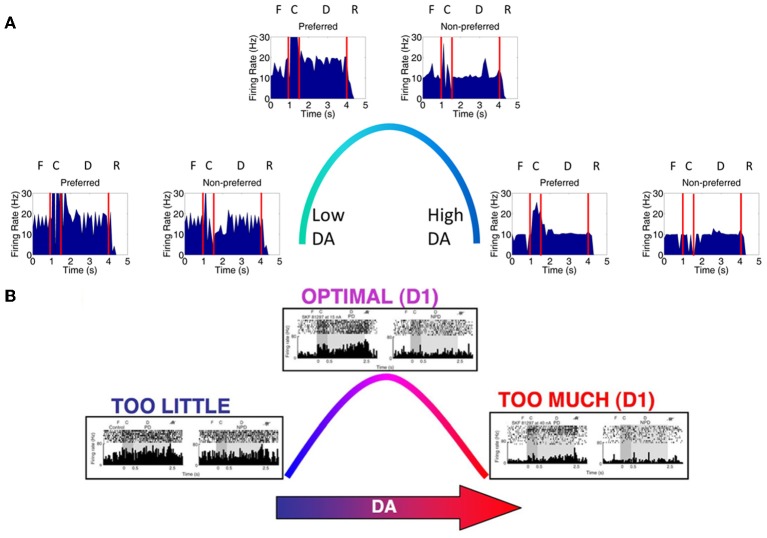
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous