

doi: 10.1128/JVI.02436-13.
Epub 2013 Oct 9.
Fatal measles virus infection prevented by brain-penetrant fusion inhibitors
Affiliations
- PMID: 24109233
- PMCID: PMC3838268
- DOI: 10.1128/JVI.02436-13
Item in Clipboard
Fatal measles virus infection prevented by brain-penetrant fusion inhibitors
J Virol.
2013 Dec.
Abstract
Measles virus (MV) infection causes an acute childhood disease that can include infection of the central nervous system and can rarely progress to severe neurological disease for which there is no specific treatment. We generated potent antiviral peptide inhibitors of MV entry and spreading and MV-induced cell fusion. Dimers of MV-specific peptides derived from the C-terminal heptad repeat region of the MV fusion protein, conjugated to cholesterol, efficiently protect SLAM transgenic mice from fatal MV infection. Fusion inhibitors hold promise for the prophylaxis of MV infection in unvaccinated and immunocompromised people, as well as potential for the treatment of grave neurological complications of measles.
Figures
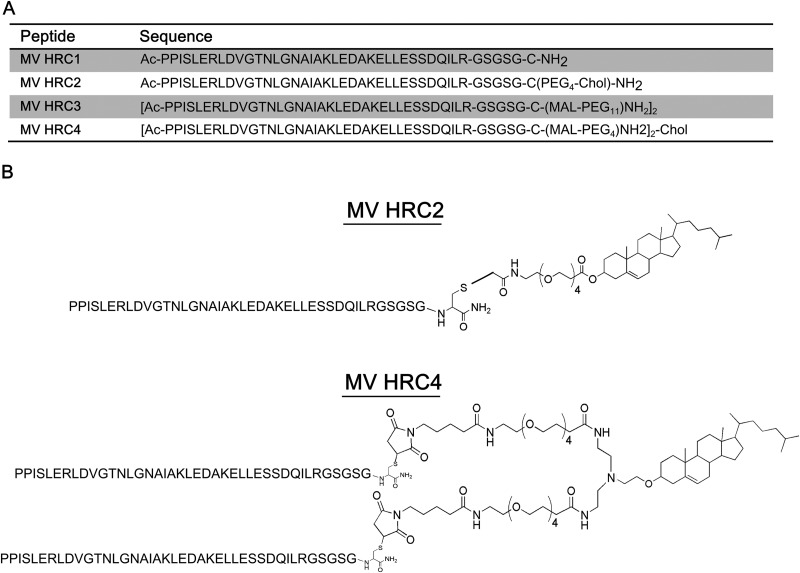
Sequences and structures of the MV HRC peptides. (A)The peptides consist of the MV HRC (amino acids 450 to 485 of MV F) with a GSGSG spacer, either dimerized (HRC3 and HRC4) or monomeric (HRC1 and HRC2). Specific peptides are linked to a PEG spacer (HRC2, HRC3, and HRC4) and modified by cholesterol (Chol) conjugation (HRC2 and HRC4). Ac, acetyl. (B) Schematic representation of MV peptides HRC2 and HRC4.
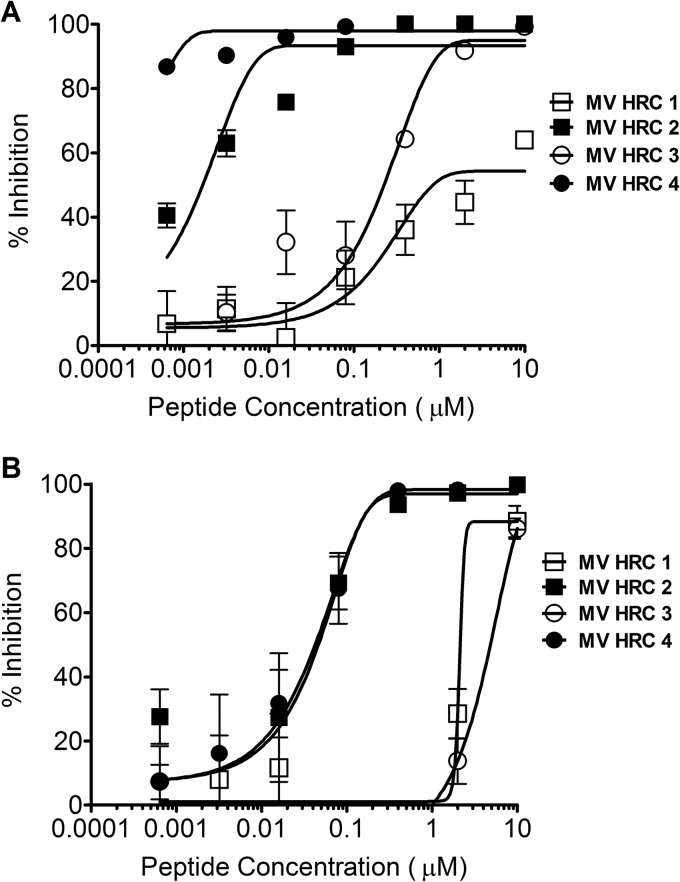
Inhibition of MV entry and spread. (A) Inhibition of MV entry by MV HRC peptides. Vero-SLAM cell monolayers were infected with WT MV G954 in the presence of the indicated concentrations of MV HRC peptides. Viral entry was assessed by plaque reduction assay (multiplicity of infection [MOI], 2 × 10−4). Results are presented as percent reductions (y axis) in the number of plaques compared to that in the absence of treatment, as a function of the compound concentration (x axis). Each point represents the mean of four experiments (± the standard error). MV HRC4 is significantly more efficient at inhibiting entry than MV HRC2 is (P = 0.0016, F test). (B) Inhibition of MV spread by MV HRC peptides. Vero-SLAM cell monolayers were infected with WT MV-IC323-EGFP. After 90 min, medium with the indicated peptide concentrations was added. Viral spread was assessed by using a readout of EGFP fluorescence after 72 h (MOI, 2 × 10−4). Results are presented as percent reductions in fluorescence (y axis) compared to that in the absence of treatment, as a function of the compound concentration (x axis). Each point represents the mean of a representative experiment (± the standard deviation).
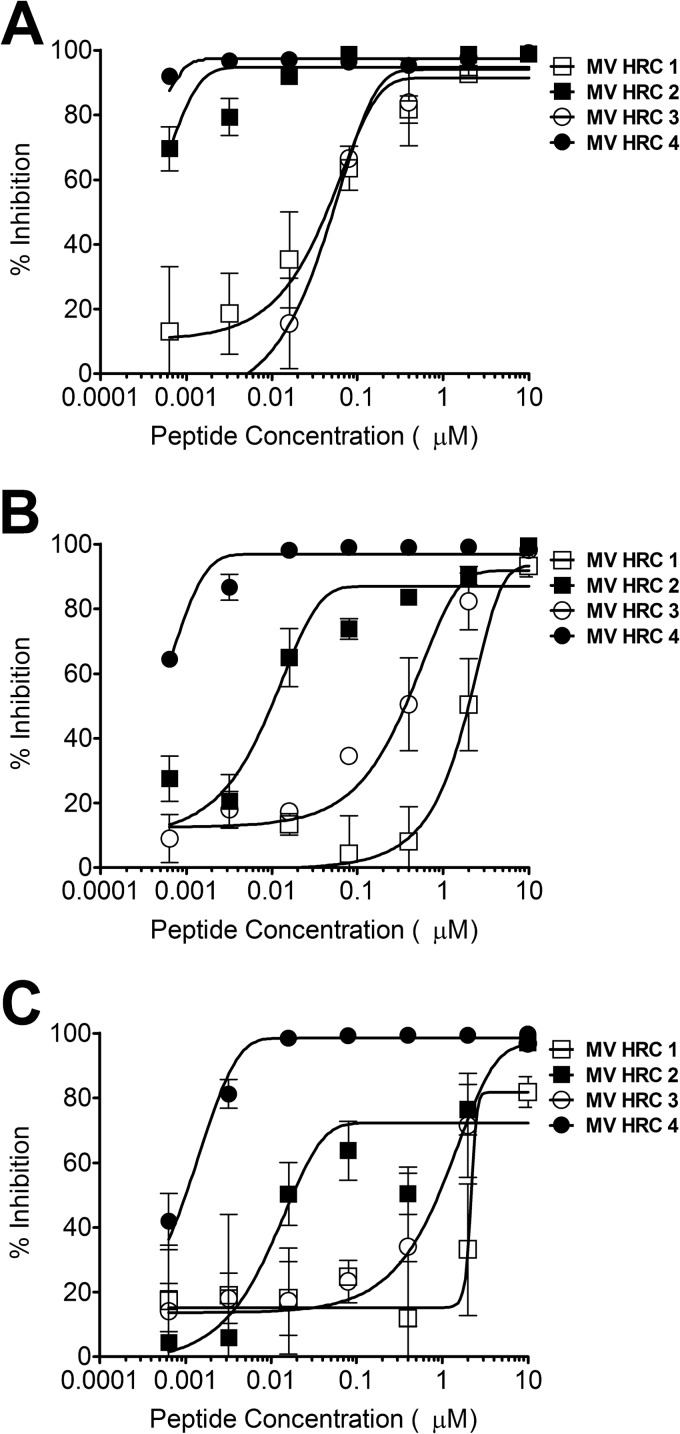
Inhibition of cell fusion. Fusion of MV H- and F-coexpressing cells with SLAM-bearing cells in the presence of the indicated compound concentrations was quantitated at 2 h (A), 4 h (B), and 6 h (C) with a β-Gal complementation assay. Results are presented as percent reductions in luminescence (y axis) compared to that in the absence of treatment, as a function of the compound concentration (x axis). Each point is the mean result of three experiments (± the standard error). MV HRC4 is significantly more efficient at inhibiting fusion than MV HRC2 is (P < 0.0001, F test).
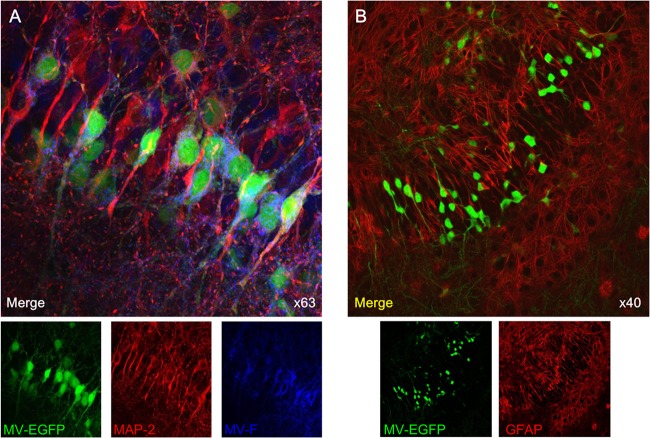
MV infection of SLAM TG organotypic brain explants. Hippocampal slices obtained from SLAM TG murine brains were infected with MV-IC323-EGFP at 104 PFU/slice of for 3 days, stained with the indicated Abs, and analyzed by confocal microscopy. (A) MV-infected slices (green) were stained with neuron-specific MAP-2 (red)- and MV F-specific (blue) Abs. (B) MV-infected slices (green for EGFP) were stained with an astrocyte-specific GFAP (red) Ab and did not show any colocalization.
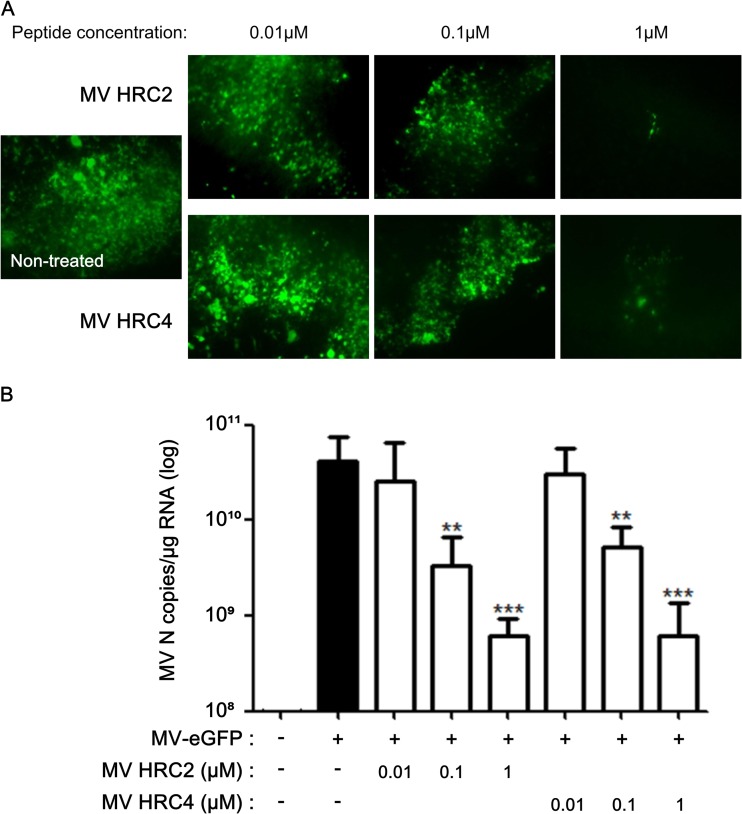
MV HRC peptides inhibit MV infection of organotypic brain explants. Hippocampal slices from SLAM TG murine brains were treated with the indicated concentrations of either HRC2 or HRC4 for 5 h or left untreated (control) and infected with MV-IC323-EGFP at 104 PFU/slice for 3 days. (A) Cultures from MV-infected explants (green) were observed by immunofluorescence microscopy. (B) Total RNA was harvested from organotypic slices at 3 days postinfection, and the level of MV N gene expression was quantified by reverse transcription-qPCR. Results are expressed as means ± standard deviations of triplicate cultures (**, P < 0.01; ***, P < 0.001 [unpaired t test]).
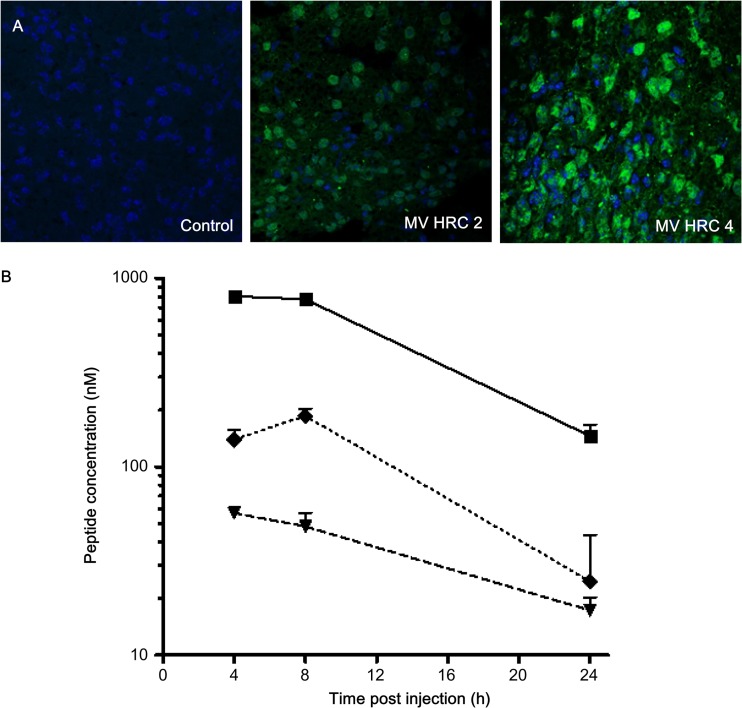
Biodistribution of MV HRC peptides. (A) Mice were injected s.c. with MV HRC2 or HRC4 or the vehicle (control) at 6 mg/kg. Animals were sacrificed 8 h later, and their brains were stained with peptide-specific Abs. (B). Mice were injected s.c. with MV HRC4 at 6 mg/kg. At the indicated times points, the animals were sacrificed and the peptide concentrations in serum (square, smooth line), brain (triangle, dashed line), and lung (diamond, dotted line) samples were quantitated. The value at each point is the mean result obtained from three animals (± the standard deviation).
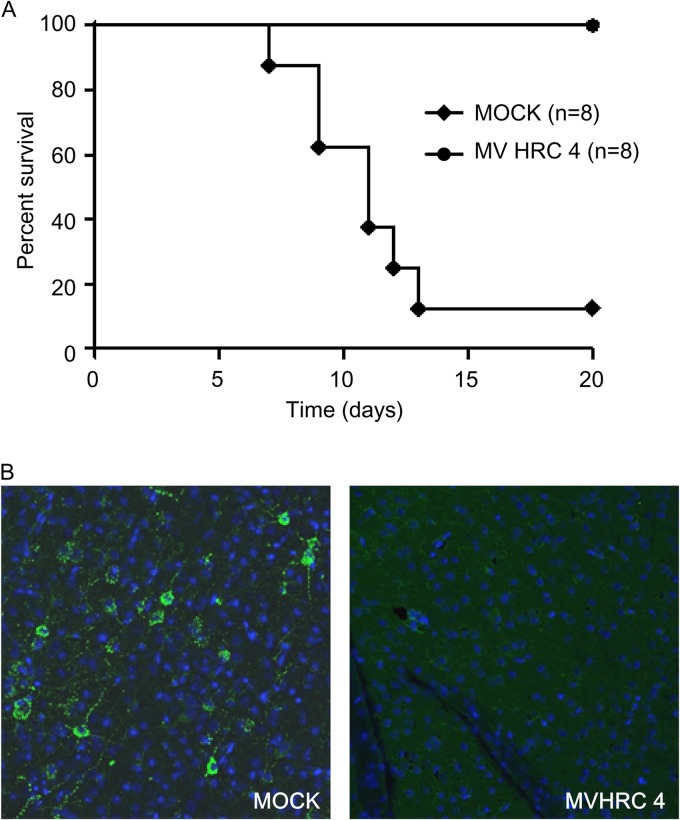
MV HRC4 protects suckling mice from lethal MV encephalitis. One-week-old SLAM TG mice received an i.n. challenge of MV G954 at 24 h after MV-HRC peptide treatment and were followed for 5 weeks postinfection. Controls animals (MOCK) were injected with the vehicle alone. (A) Mice received prophylactic i.n. administration of peptides, followed by s.c. injection after infection, daily for 7 days. The statistical significance of the difference between the MV HRC4- and mock-injected groups was analyzed with the Mantel-Cox test (P = 0.0004). (B) Immunofluorescence analysis of MV N expression in the brain of either mock-treated animals on the day that neurological signs appeared or surviving MV HRC4-treated animals at the end of the experiment. Representative images from three mice analyzed in each group are presented (magnification, ×40).
References
-
- Simons E, Ferrari M, Fricks J, Wannemuehler K, Anand A, Burton A, Strebel P. 2012. Assessment of the 2010 global measles mortality reduction goal: results from a model of surveillance data. Lancet 379:2173–2178 - PubMed
-
- Moss WJ, Griffin DE. 2012. Measles. Lancet 379:153–164 - PubMed
-
- De Serres G, Markowski F, Toth E, Landry M, Auger D, Mercier M, Belanger P, Turmel B, Arruda H, Boulianne N, Ward BJ, Skowronski DM. 2013. The largest measles epidemic in North America in a decade—Quebec, Canada, 2011: contribution of susceptibility, serendipity and super-spreading events on elimination. J. Infect. Dis. 207:990–998 - PubMed
-
- Hosoya M. 2006. Measles encephalitis: direct viral invasion or autoimmune-mediated inflammation? Intern. Med. 45:841–842 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
