

Rapid effects of oestrogen on synaptic plasticity: interactions with actin and its signalling proteins
- PMID: 24112361
- PMCID: PMC3989941
- DOI: 10.1111/jne.12108
Rapid effects of oestrogen on synaptic plasticity: interactions with actin and its signalling proteins
Abstract
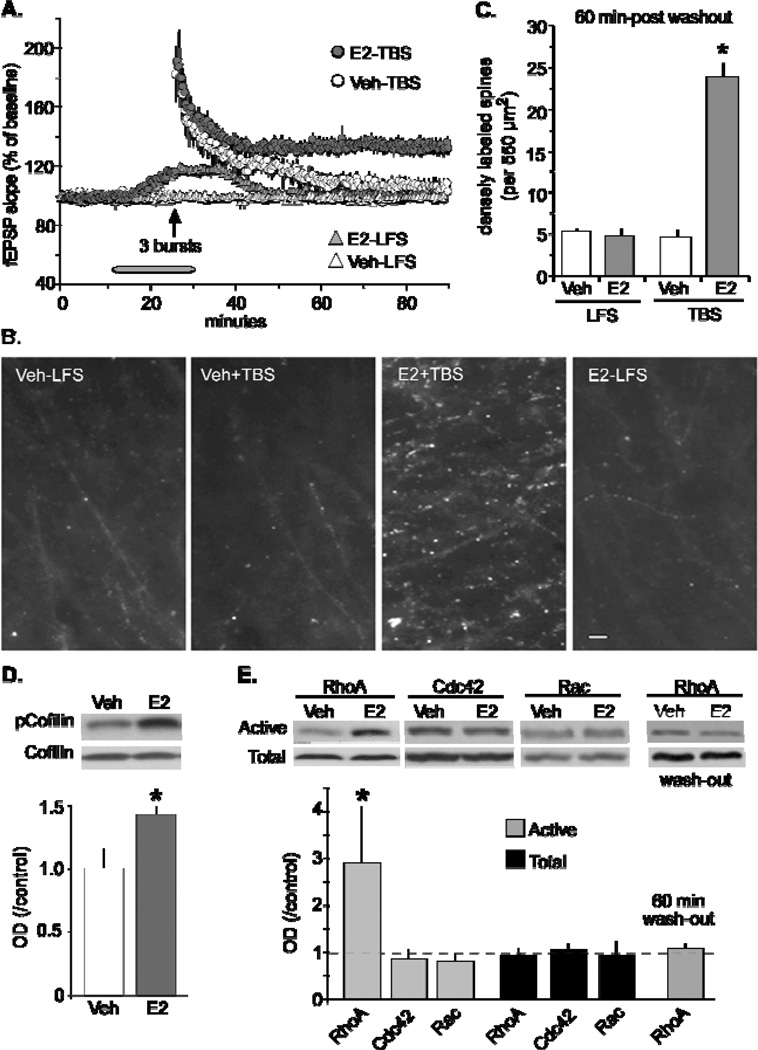
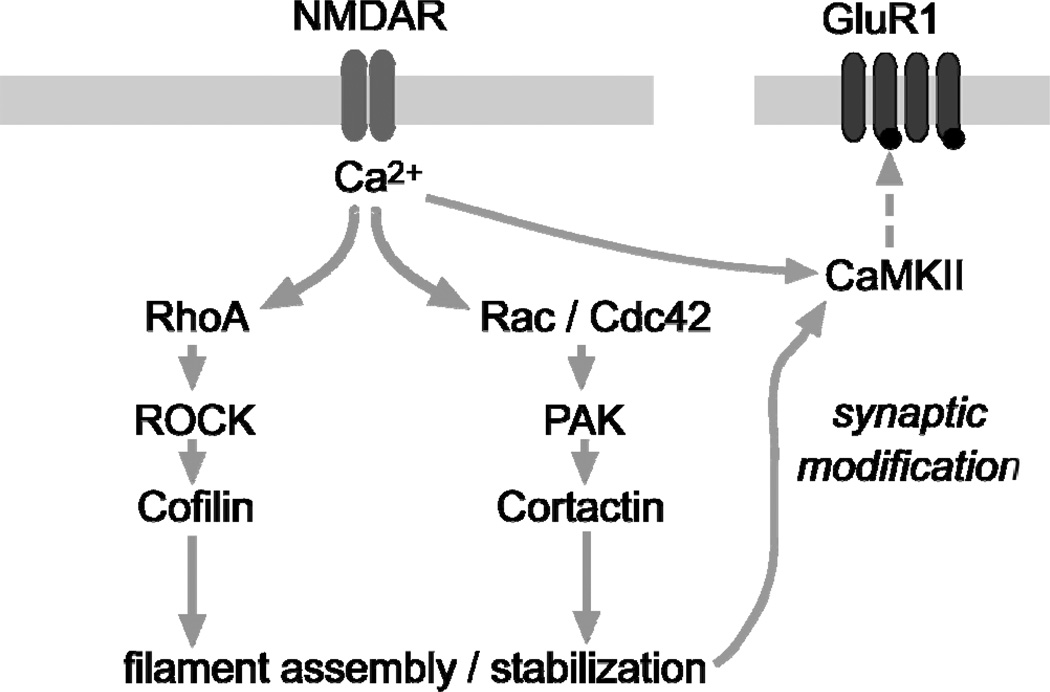
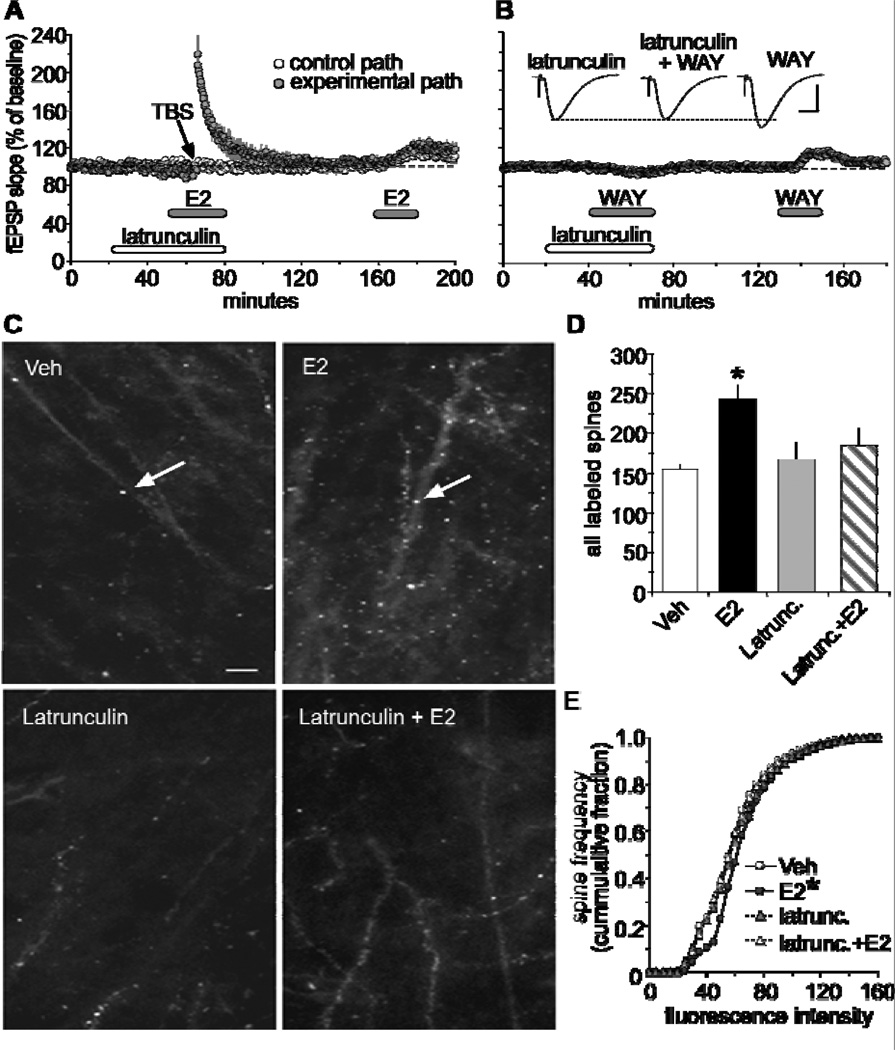
Oestrogen rapidly enhances fast excitatory postsynaptic potentials, facilitates long-term potentiation (LTP) and increases spine numbers. Each effect likely contributes to the influence of the steroid on cognition and memory. In the present review, we first describe a model for the substrates of LTP that includes an outline of the synaptic events occurring during induction, expression and consolidation. Briefly, critical signalling pathways involving the small GTPases RhoA and Rac/Cdc42 are activated by theta burst-induced calcium influx and initiate actin filament assembly via phosphorylation (inactivation) of cofilin. Reorganisation of the actin cytoskeleton changes spine and synapse morphology, resulting in increased concentrations of AMPA receptors at stimulated contacts. We then use the synaptic model to develop a specific hypothesis about how oestrogen affects both baseline transmission and plasticity. Brief infusions of 17β-oestradiol (E2 ) reversibly stimulate the RhoA, cofilin phosphorylation and actin polymerisation cascade of the LTP machinery; blocking this eliminates the effects of the steroid on transmission. We accordingly propose that E2 induces a weak form of LTP and thereby increases synaptic responses, a hypothesis that also accounts for how it markedly enhances theta burst induced potentiation. Although the effects of E2 on the cytoskeleton could be a result of the direct activation of small GTPases by oestrogen receptors on the synaptic membrane, the hormone also activates tropomyosin-related kinase B receptors for brain-derived neurotrophic factor, a neurotrophin that engages the RhoA-cofilin sequence and promotes LTP. The latter observations raise the possibility that E2 produces its effects on synaptic physiology via transactivation of neighbouring receptors that have prominent roles in the management of spine actin, synaptic physiology and plasticity.
Keywords: RhoA; cofilin; long-term potentiation; oestradiol; pTrkB; spines.
© 2013 British Society for Neuroendocrinology.
Figures
References
-
- Devi G, Hahn K, Massimi S, Zhivotovskaya E. Prevalence of memory loss complaints and other symptoms associated with the menopause transition: a community survey. Gender medicine. 2005;2:255–264. - PubMed
-
- Wegesin DJ, Stern Y. Effects of hormone replacement therapy and aging on cognition: evidence for executive dysfunction. Neuropsychology, development, and cognition. Section B, Aging, neuropsychology and cognition. 2007;14:301–328. - PubMed
-
- Loy R, Gerlach JL, McEwen BS. Autoradiographic localization of estradiol-binding neurons in the rat hippocampal formation and entorhinal cortex. Brain Res. 1988;467:245–251. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
