

Plant genome editing made easy: targeted mutagenesis in model and crop plants using the CRISPR/Cas system
- PMID: 24112467
- PMCID: PMC3852272
- DOI: 10.1186/1746-4811-9-39
Plant genome editing made easy: targeted mutagenesis in model and crop plants using the CRISPR/Cas system
Abstract
Targeted genome engineering (also known as genome editing) has emerged as an alternative to classical plant breeding and transgenic (GMO) methods to improve crop plants. Until recently, available tools for introducing site-specific double strand DNA breaks were restricted to zinc finger nucleases (ZFNs) and TAL effector nucleases (TALENs). However, these technologies have not been widely adopted by the plant research community due to complicated design and laborious assembly of specific DNA binding proteins for each target gene. Recently, an easier method has emerged based on the bacterial type II CRISPR (clustered regularly interspaced short palindromic repeats)/Cas (CRISPR-associated) immune system. The CRISPR/Cas system allows targeted cleavage of genomic DNA guided by a customizable small noncoding RNA, resulting in gene modifications by both non-homologous end joining (NHEJ) and homology-directed repair (HDR) mechanisms. In this review we summarize and discuss recent applications of the CRISPR/Cas technology in plants.
Figures
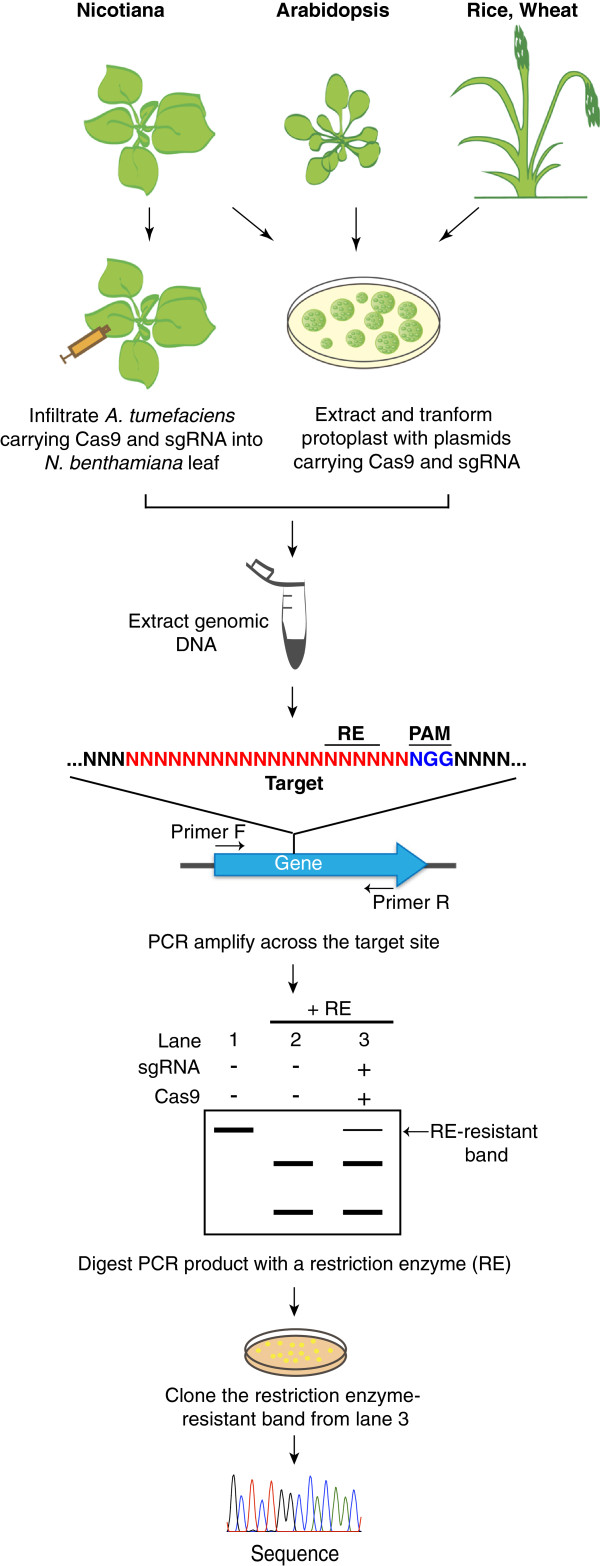
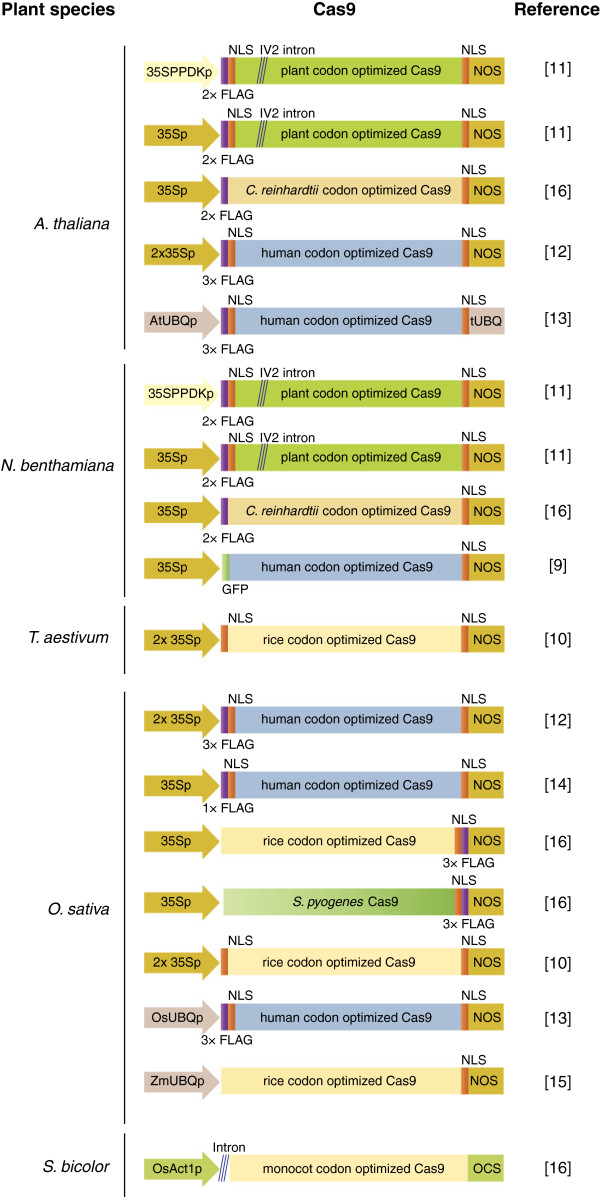
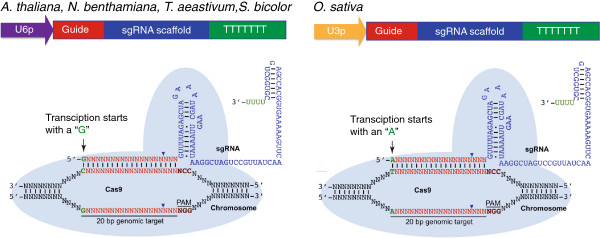
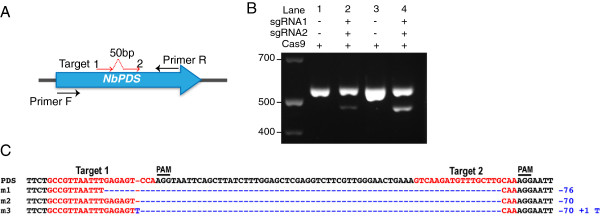
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
