

Flow cytometry-based enrichment for cell shape mutants identifies multiple genes that influence Helicobacter pylori morphology
- PMID: 24112477
- PMCID: PMC3844677
- DOI: 10.1111/mmi.12405
Flow cytometry-based enrichment for cell shape mutants identifies multiple genes that influence Helicobacter pylori morphology
Abstract
The helical cell shape of Helicobacter pylori is highly conserved and contributes to its ability to swim through and colonize the viscous gastric mucus layer. A multi-faceted peptidoglycan (PG) modification programme involving four recently characterized peptidases and two accessory proteins is essential for maintaining H. pylori's helicity. To expedite identification of additional shape-determining genes, we employed flow cytometry with fluorescence-activated cell sorting (FACS) to enrich a transposon library for bacterial cells with altered light scattering profiles that correlate with perturbed cell morphology. After a single round of sorting, 15% of our clones exhibited a stable cell shape defect, reflecting 37-fold enrichment. Sorted clones with straight rod morphology contained insertions in known PG peptidases, as well as an insertion in csd6, which we demonstrated has ld-carboxypeptidase activity and cleaves monomeric tetrapeptides in the PG sacculus, yielding tripeptides. Other mutants had only slight changes in helicity due to insertions in genes encoding MviN/MurJ, a protein possibly involved in initiating PG synthesis, and the hypothetical protein HPG27_782. Our findings demonstrate FACS robustly detects perturbations of bacterial cell shape and identify additional PG peptide modifications associated with helical cell shape in H. pylori.
© 2013 John Wiley & Sons Ltd.
Figures
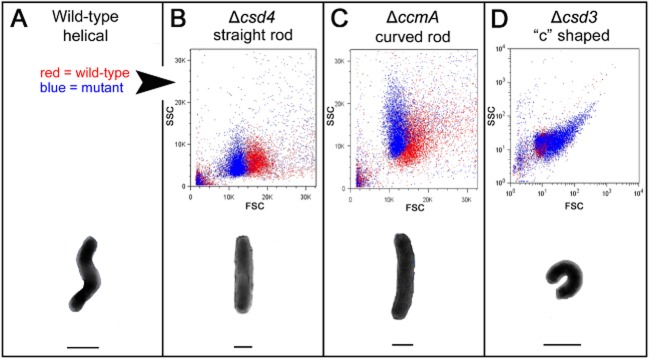
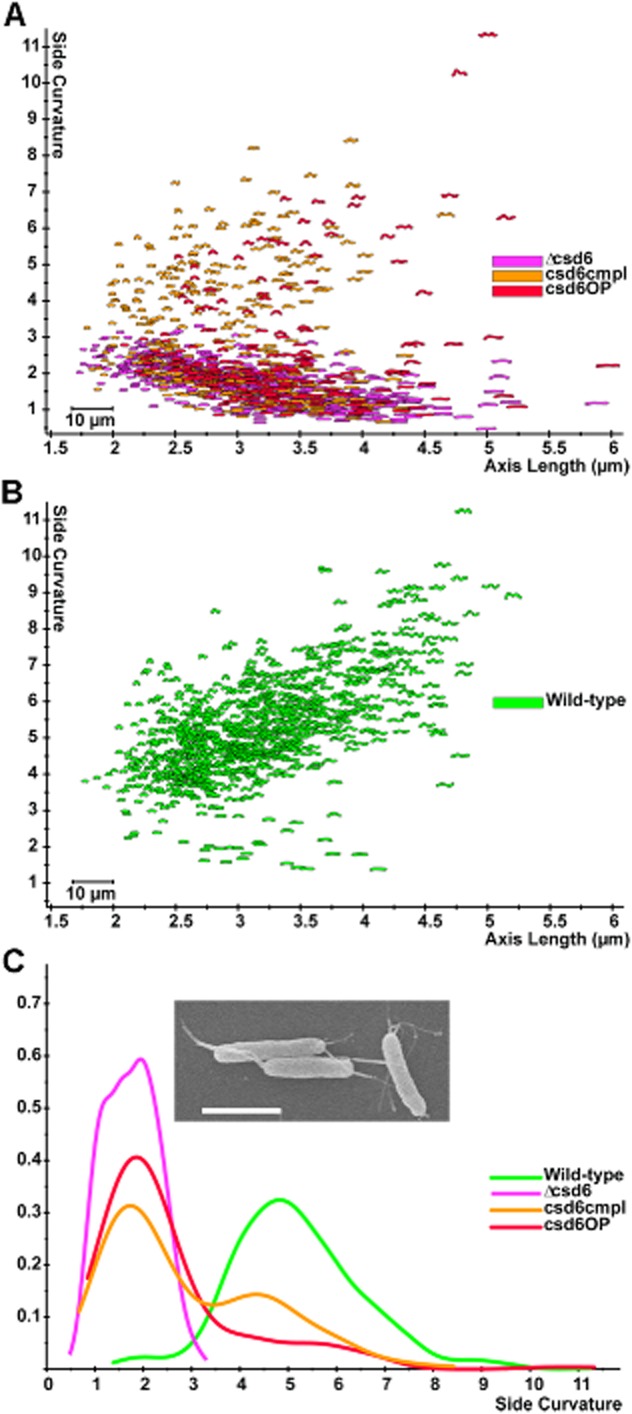
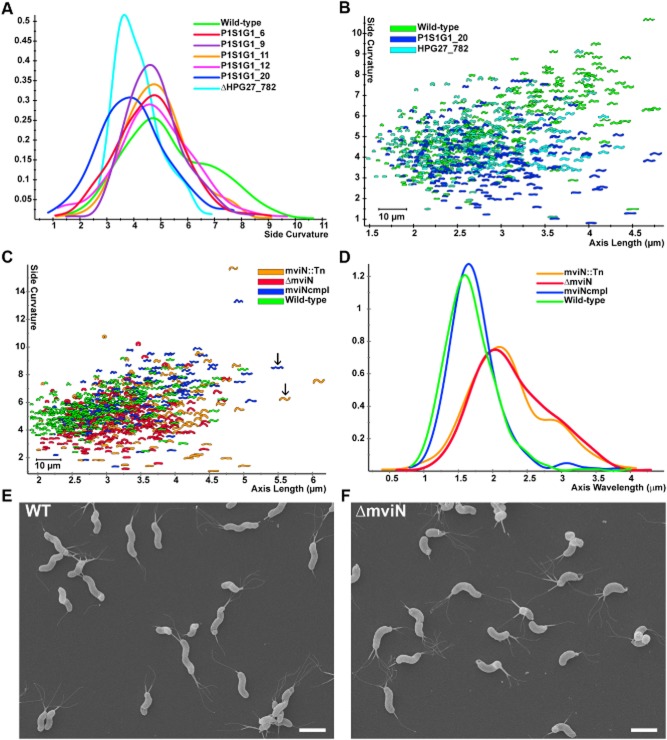
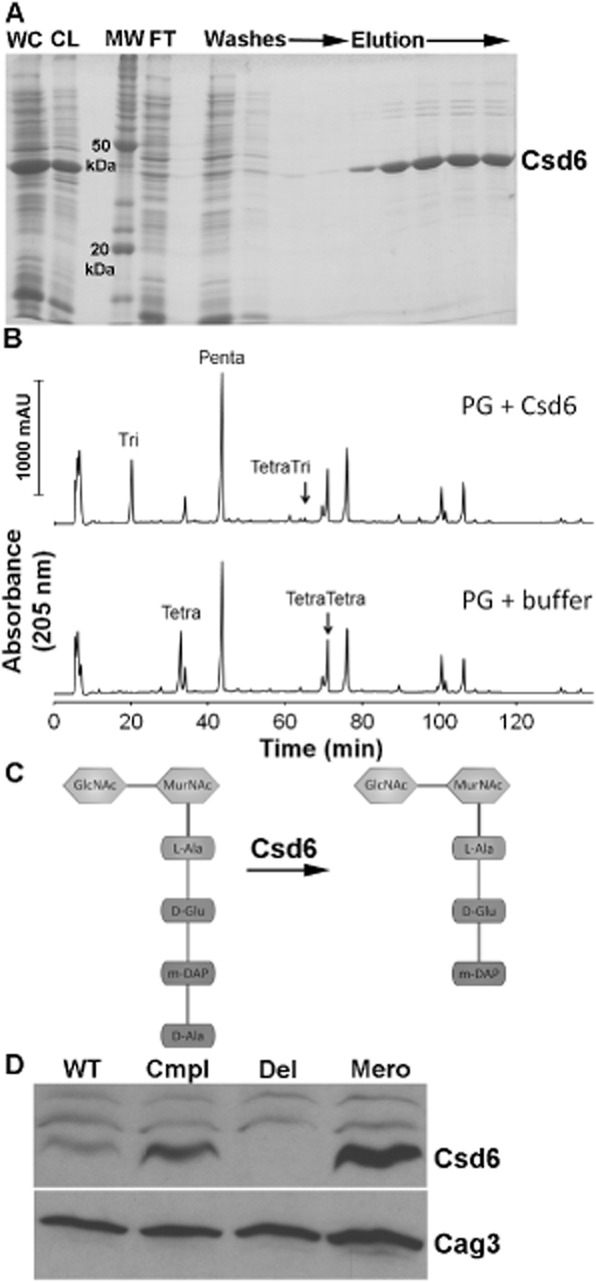
References
-
- Asakura H, Churin Y, Bauer B, Boettcher JP, Bartfeld S, Hashii N, et al. Helicobacter pylori HP0518 affects flagellin glycosylation to alter bacterial motility. Mol Microbiol. 2010;78:1130–1144. - PubMed
-
- Ausmees N, Kuhn JR. Jacobs-Wagner C. The bacterial cytoskeleton: an intermediate filament-like function in cell shape. Cell. 2003;115:705–713. - PubMed
-
- Ausubel F, Brent R, Kingston RE, Moore DD, Seidman JG, Smith JA. Struhl K. Short Protocols in Molecular Biology. New York: John Wiley & Sons; 1997.
-
- Biarrotte-Sorin S, Hugonnet JE, Delfosse V, Mainardi JL, Gutmann L, Arthur M. Mayer C. Crystal structure of a novel beta-lactam-insensitive peptidoglycan transpeptidase. J Mol Biol. 2006;359:533–538. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
