

Review
doi: 10.1038/nri3536.
Epub 2013 Oct 11.
T-bet: a bridge between innate and adaptive immunity
Affiliations
- PMID: 24113868
- PMCID: PMC6290922
- DOI: 10.1038/nri3536
Item in Clipboard
Review
T-bet: a bridge between innate and adaptive immunity
Nat Rev Immunol.
2013 Nov.
Abstract
Originally described over a decade ago as a T cell transcription factor regulating T helper 1 cell lineage commitment, T-bet is now recognized as having an important role in many cells of the adaptive and innate immune system. T-bet has a fundamental role in coordinating type 1 immune responses by controlling a network of genetic programmes that regulate the development of certain immune cells and the effector functions of others. Many of these transcriptional networks are conserved across innate and adaptive immune cells and these shared mechanisms highlight the biological functions that are regulated by T-bet.
Conflict of interest statement
Competing interests statement
The authors declare
Figures
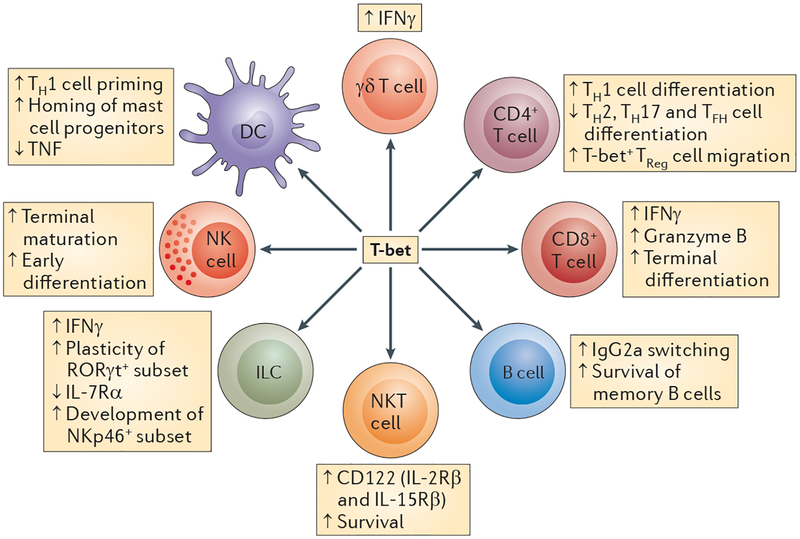
T-bet is expressed in multiple cells of the innate and adaptive immune system. Its expression is required for the survival, development and proper functions of immune cells. In the innate immune system, T-bet is expressed in dendritic cells (DCs), natural killer (NK) cells, natural killer T (NKT) cells and innate lymphoid cells (ILCs). In the adaptive immune system, T-bet is expressed in CD4+ and CD8+ T effector cells, B cells, γδ T cells and a subset of regulatory T (TReg) cells. IFNγ, interferon-γ; IL-7Rα, interleukin-7 receptor-α; NKp46, NK cell p46-related protein; RORγt, retinoic acid receptor-related orphan receptor-γt; TFH, T follicular helper; TH, T helper; TNF, tumour necrosis factor.
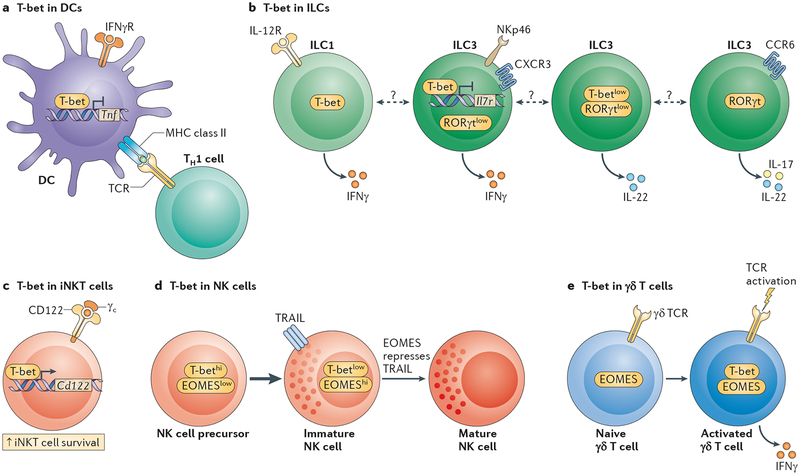
a | T-bet expression in dendritic cells (DCs) is required to properly prime T helper 1 (TH1) cells. T-bet suppresses tumour necrosis factor (TNF) production in colonic DCs and this is required for the maintenance of mucosal homeostasis. b | T-bet is expressed in the innate lymphoid cell 1 (ILC1) subset, which is characterized by the sole production of interferon-γ (IFNγ). ILC3s are dependent on the expression of retinoic acid receptor-related orphan receptor-γt (RORγt) and can be subdivided into CC-chemokine receptor 6 (CCR6)+ and CCR6– subsets. CCR6− ILC3s express T-bet. High T-bet expression in these cells is associated with low RORγt and interleukin-7 receptor (IL-7R) expression, and high expression of NK cell p46-related protein (NKp46), CXC-chemokine receptor 3 (CXCR3) and IFNγ. Low T-bet expression in the CCR6− ILC3 subset is associated with IL-22 expression and low or no expression of NKp46 and IFNγ. CCR6+ ILC3s do not express T-bet but express IL-17 and IL-22. The lineage inter-relationships of these different subpopulations are incompletely defined. c | T-bet expression in invariant natural killer T (iNKT) cells promotes their survival through the regulation of CD122 (also known as IL-15Rβ) expression. d | T-bet and eomesodermin (EOMES) regulate the maturation process of NK cells in a coordinated fashion. T-bet is expressed at an immature stage of differentiation that is characterized by TNF-related apoptosis-inducing ligand (TRAIL) expression. EOMES expression is required to silence TRAIL expression and to complete the maturation process of NK cells. e | T-bet is not expressed in naive γδ T cells. Its expression is rapidly induced following T cell receptor (TCR) engagement. Together with EOMES, T-bet regulates IFNγ production in mature γδ T cells. Dashed line indicates that the developmental relationship between these cells is unclear. γc, common cytokine receptor γ-chain; IFNγR, IFNγ receptor.
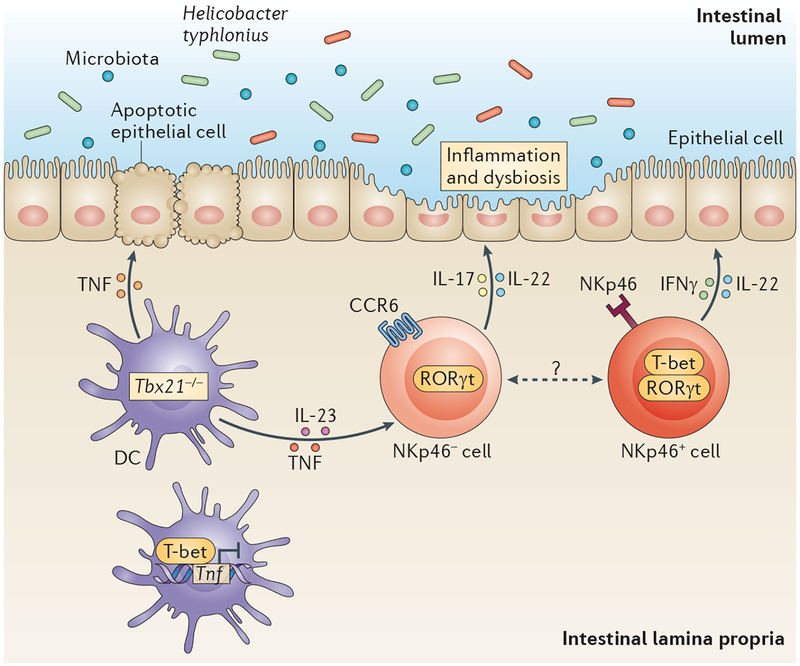
Innate lymphoid cells (ILCs) and T-bet-expressing dendritic cells (DCs) interact in the colonic lamina propria to maintain epithelial integrity. T-bet (encoded by Tbx21) represses tumour necrosis factor (TNF) expression in colonic DCs. In the absence of T-bet, TNF from DCs causes colonic epithelial cell apoptosis in the context of the pathobiont, Helicobacter typhlonius. TNF from DCs cooperates with interleukin-23 (IL-23) to induce interferon-γ (IFNγ) and IL-17 expression from ILCs. T-bet expression in ILCs regulates the plasticity of cytokine responses by repressing IL-17 and by inducing IFNγ production. This pathology creates an inflammatory milieu that generates a dysbiotic microbiota that is capable of transmitting colonic inflammation to wild-type mice. Dashed line indicates that the developmental relationship between these ILC subsets is unclear. CCR6, CC-chemokine receptor 6; NKp46, NK cell p46-related protein; RORγt, retinoic acid receptor-related orphan receptor-γt.
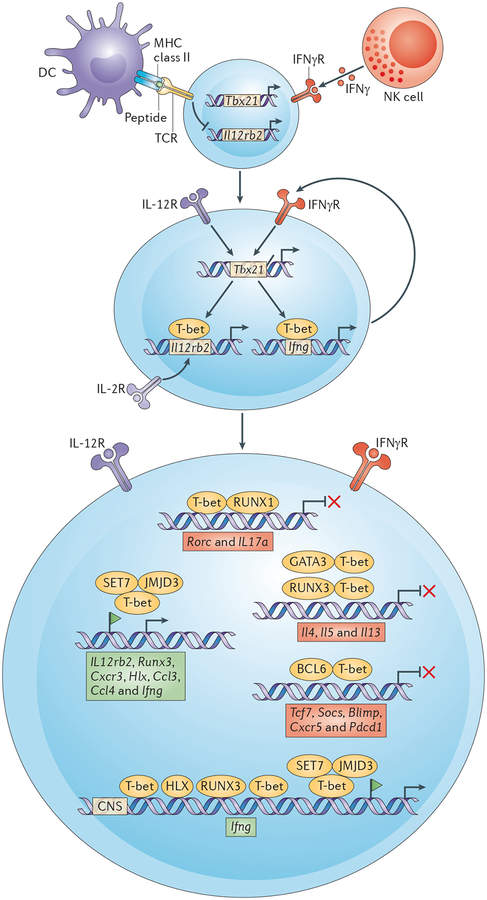
T cell receptor (TCR) and interferon-γ (IFNγ)-mediated signalling induces the first wave of T-bet (encoded by Tbx21) expression in an interleukin-12 (IL-12)independent manner, as signalling through TCR inhibits the expression of IL-12 receptor-β2 (Il12rb2) subunit. Cessation of TCR stimulation and IL-2 signalling induces expression of the IL-12Rβ2 subunit, which thus enables IL-12 signalling. IL-12-mediated activation of signal transducer and activator of transcription 4 (STAT4) prompts the second wave of T-bet expression. IFNγ produced by T helper 1 (TH1) cells functions in a feedforward loop, which further increases T-bet expression in TH1 cells. T-bet recruits enzymes (jumonji domain-containing protein 3 (JMJD3) and SET domain-containing protein 7 (SET7; also known as SET9)), which generate chromatin modifications that are associated with gene activation. T-bet-mediated transactivation of the Ifng gene is enhanced by H2.0-like homeobox protein 1 (HLX) and runt-related transcription factor 3 (RUNX3) transcription factors, both of which are encoded by T-bet target genes. T-bet facilitates CCCTC-binding factor (CTCF) binding and chromatin looping at the Ifng locus, which is required for optimal IFNγ expression in TH1 cells. T-bet induces chemokines, chemokine receptors and other effector molecules that are required for proper migration and function of TH1 cells. Sequestration of GATA-binding protein 3 (GATA3) by T-bet prevents the GATA3-mediated activation of the Il4–Il5–Il13 locus, and binding of a T-bet–RUNX3 complex to the Il4 silencer (DNase I hypersensitivity site 4 (HS4)) prevents the expression of Il4 in TH precursor cells. T-bet also binds to the Gata3 locus, and its binding is associated with the presence of repressive epigenetic marks. RUNX1 enhances the expression of retinoic acid receptor-related orphan receptor-γt (RORγt; encoded by Rorc) in TH17 cells and it functions as a RORγt-specific co-activator of the Il17a gene. T-bet binding to RUNX1 prevents Rorc and Il17a expression in TH precursor cells. Furthermore, T-bet together with STAT4, silences the Rorc locus. T-bet interacts with B cell lymphoma 6 (BCL-6) and recruits it to the promoters of genes that are repressed by T-bet in TH1 cells (for example, T cell factor (Tcf7) and suppressor of cytokine signalling (Socs)). By sequestering BCL-6 away from BCL-6 target genes (for example B lymphocyte-induced maturation protein (Blimp), CXC-chemokine receptor 5 (Cxcr5) and programmed cell death 1 (Pdcd1), T-bet effectively blocks T follicular helper (TFH) cell lineage commitment. Ccl, CC-chemokine ligand; CNS, conserved non-coding sequence; DC, dendritic cell; IFNγR, IFNγ receptor; NK, natural killer.
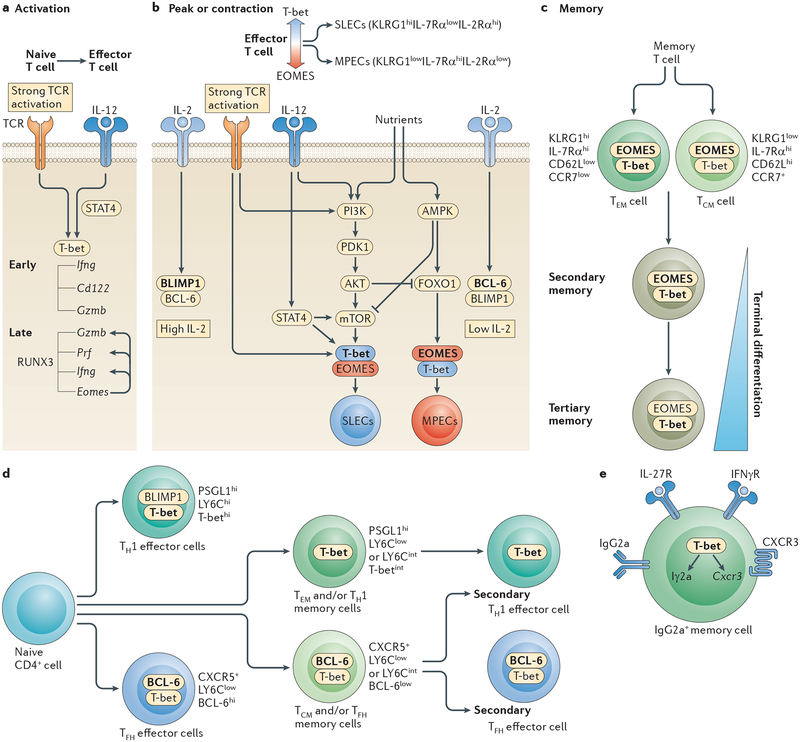
a | In CD8+ T cells, T-bet (encoded by Tbx21) expression is induced by T cell receptor (TCR) and interleukin-12 receptor (IL-12R) signalling, whereas eomesodermin (EOMES) is upregulated later in a runt-related transcription factor 3 (RUNX3)-dependent manner. RUNX3, T-bet and EOMES regulate the expression of interferon-γ (IFNγ), granzyme B (encoded by Gzmb), perforin (encoded by Prf) and CD122 in CD8+ effector cells. b | Strong TCR and IL-12R signalling promotes the terminal differentiation of short-lived effector cells (SLECs) by enhancing and maintaining the activity of the kinase mammalian target of rapamycin (mTOR) downstream of signal transducer and activator of transcription 4 (STAT4). mTOR simultaneously induces T-bet and represses EOMES expression through inhibition of forkhead box O1 (FOXO1). The IL-2R–STAT5 pathway induces the expression of B lymphocyte-induced maturation protein 1 (BLIMP1), inhibiting the expression of B cell lymphoma 6 (BCL-6), which is a transcriptional repressor of the Tbx21 gene. Inhibition of mTOR activity by rapamycin or by activators of AMP-activated protein kinase (AMPK), such as metformin, supports the development of memory precursor effector cells (MPECs). Similarly, enhanced MPEC formation is observed in low IL-2 and IL-12 conditions. c | EOMES expression increases and T-bet levels decline in memory CD8+ T cells. Minimal levels of T-bet are maintained for proper expression of CD122, which promotes the homeostatic proliferation of memory cells. T-bet expression is associated with a more differentiated phenotype of effector memory T (TEM) cells and secondary and tertiary memory cells. d | P-selectin glycoprotein ligand 1 (PSGL1)hi LY6ChiT-bethi cells are terminally differentiated T helper 1 (TH1) effector cells. PSGL1hiLY6ClowT-betint cells are less differentiated, are longer lived and give rise to TH1 effector cells after re-challenge. T-bet is expressed at very low levels in T follicular helper (TFH) and TFH memory cells, as a result of BCL-6 expression. e | T-bet is a selective inducer of IFNγ-mediated IgG2a class-switching in B cells. T-bet is required for the survival of IgG2a+CD38hi memory B cells and it promotes the migration of IgG2a+ B cells to inflammatory foci through the transcriptional regulation of CXC-chemokine receptor 3 (CXCR3) expression. CCR7; CC-chemokine receptor 7; IFNγR, IFNγ receptor; KLRG1; killer cell lectin-like receptor subfamily G member 1; PDK1, 3-phosphoinositide-dependent protein kinase 1; PI3K, phosphoinositide 3-kinase; TCM, central memory.
References
-
- Horton AC & Gibson-Brown JJ Evolution of developmental functions by the Eomesodermin, T-brain-1, Tbx21 subfamily of T-box genes: insights from amphioxus. J. Exp. Zool 294, 112–121 (2002). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
