

Poxviral protein A52 stimulates p38 mitogen-activated protein kinase (MAPK) activation by causing tumor necrosis factor receptor-associated factor 6 (TRAF6) self-association leading to transforming growth factor β-activated kinase 1 (TAK1) recruitment
- PMID: 24114841
- PMCID: PMC3837111
- DOI: 10.1074/jbc.M113.485490
Poxviral protein A52 stimulates p38 mitogen-activated protein kinase (MAPK) activation by causing tumor necrosis factor receptor-associated factor 6 (TRAF6) self-association leading to transforming growth factor β-activated kinase 1 (TAK1) recruitment
Abstract
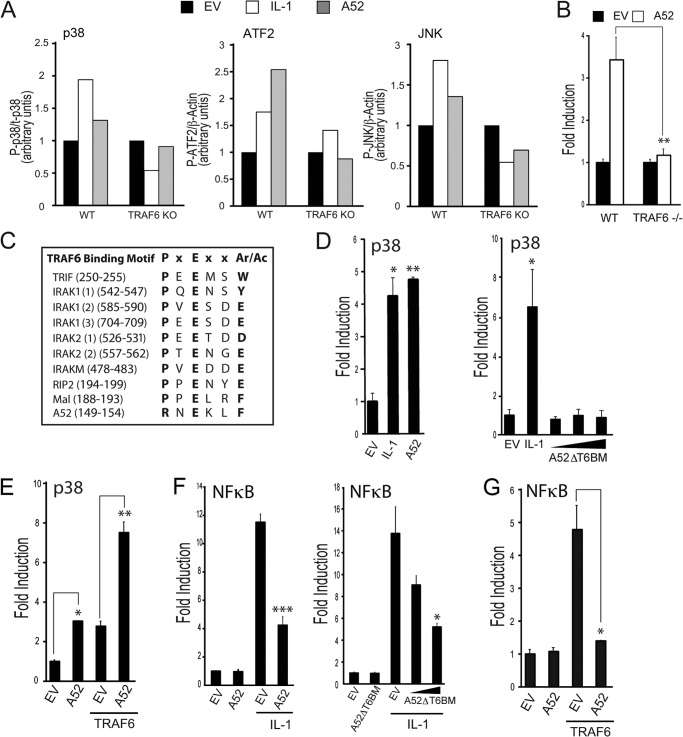
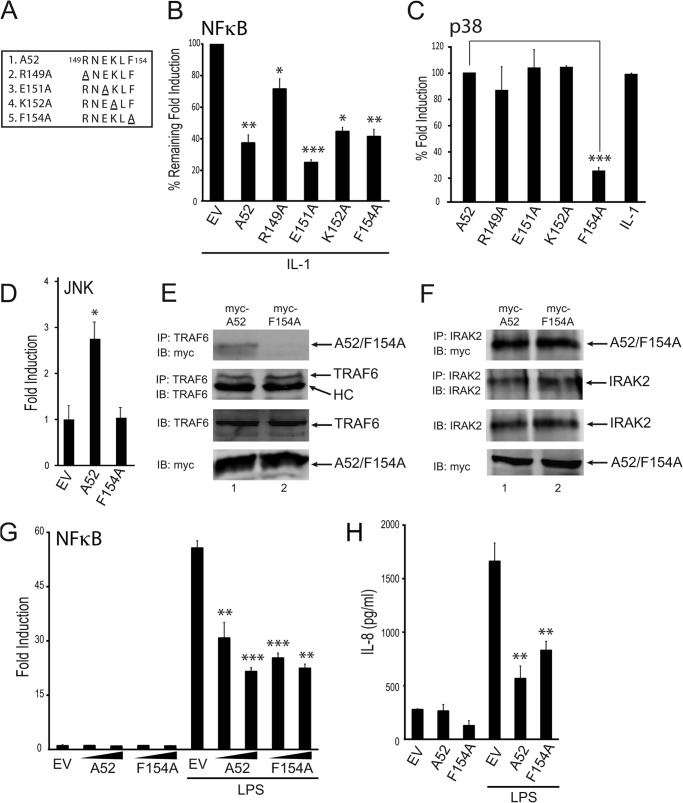
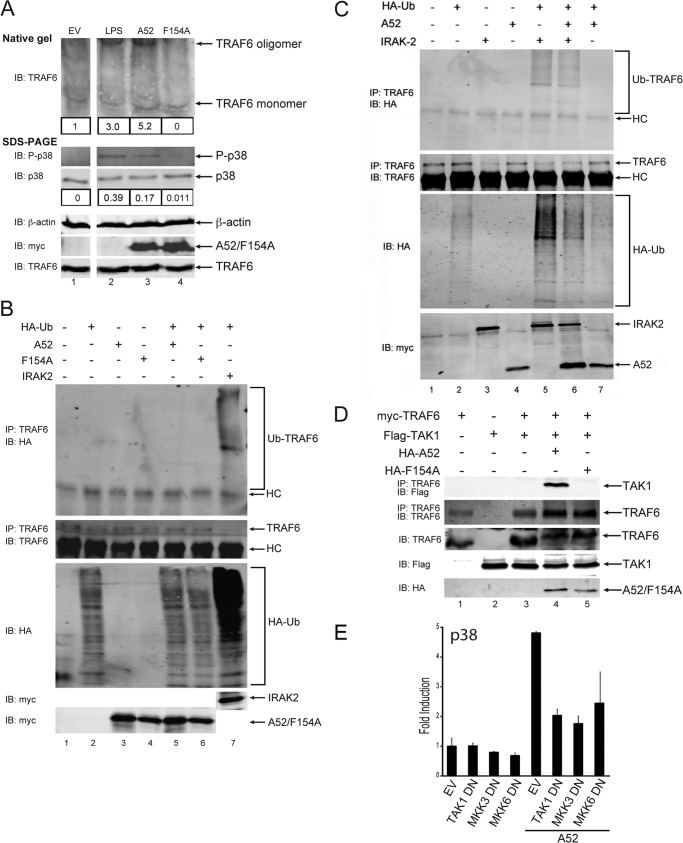
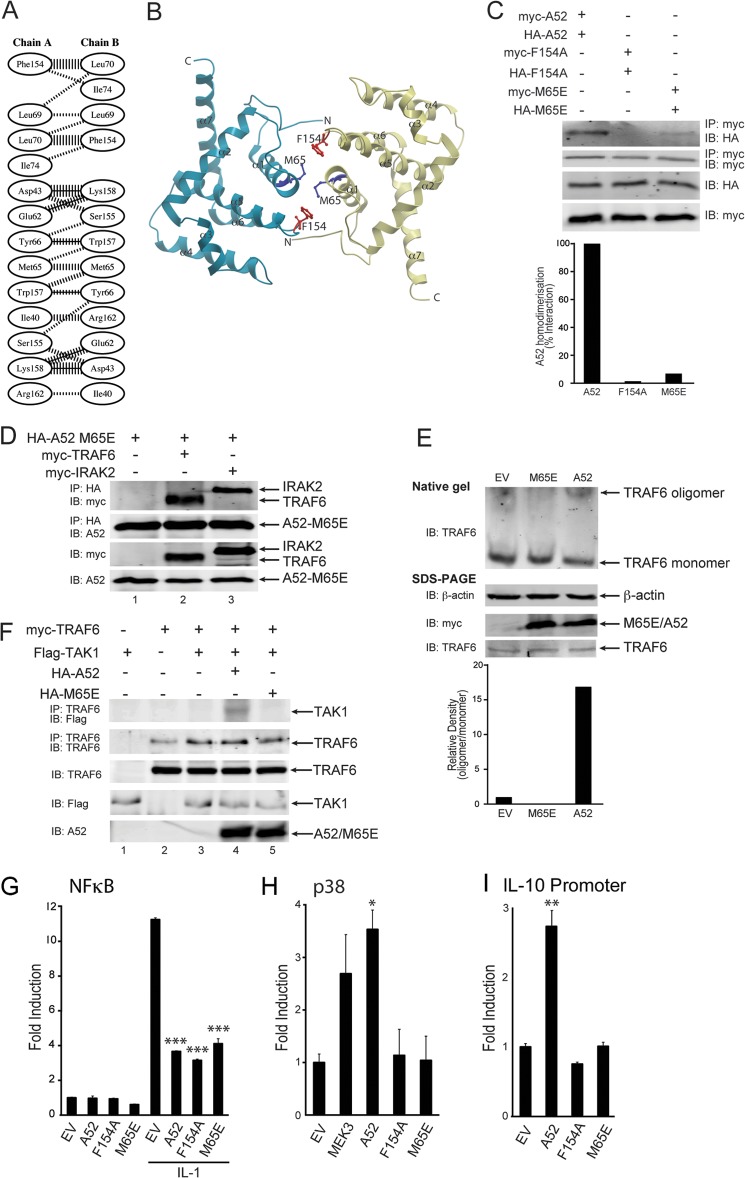
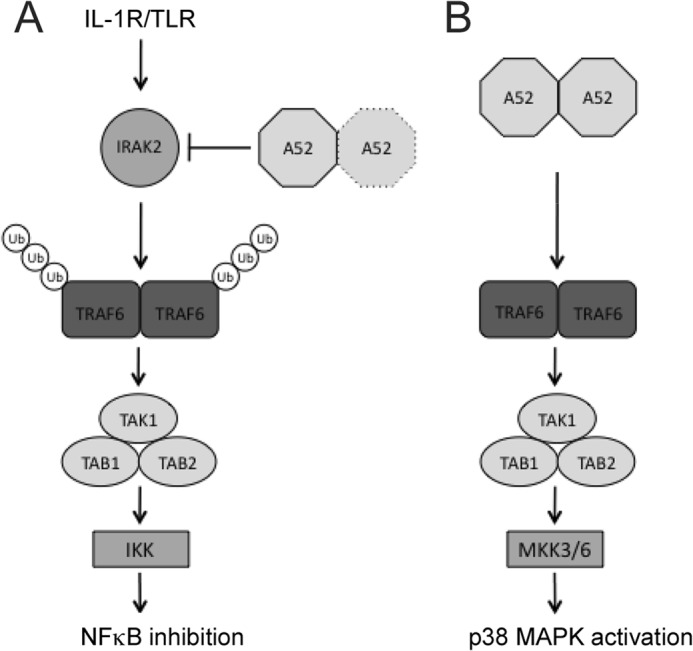
Vaccinia virus encodes a number of proteins that inhibit and manipulate innate immune signaling pathways that also have a role in virulence. These include A52, a protein shown to inhibit IL-1- and Toll-like receptor-stimulated NFκB activation, via interaction with interleukin-1 receptor-associated kinase 2 (IRAK2). Interestingly, A52 was also found to activate p38 MAPK and thus enhance Toll-like receptor-dependent IL-10 induction, which was TRAF6-dependent, but the manner in which A52 manipulates TRAF6 to stimulate p38 activation was unclear. Here, we show that A52 has a non-canonical TRAF6-binding motif that is essential for TRAF6 binding and p38 activation but dispensable for NFκB inhibition and IRAK2 interaction. Wild-type A52, but not a mutant defective in p38 activation and TRAF6 binding (F154A), caused TRAF6 oligomerization and subsequent TRAF6-TAK1 association. The crystal structure of A52 shows that it adopts a Bcl2-like fold and exists as a dimer in solution. Residue Met-65 was identified as being located in the A52 dimer interface, and consistent with that, A52-M65E was impaired in its ability to dimerize. A52-M65E although capable of interacting with TRAF6, was unable to cause either TRAF6 self-association, induce the TRAF6-TAK1 association, or activate p38 MAPK. The results suggest that an A52 dimer causes TRAF6 self-association, leading to TAK1 recruitment and p38 activation. This reveals a molecular mechanism whereby poxviruses manipulate TRAF6 to activate MAPKs (which can be proviral) without stimulating antiviral NFκB activation.
Keywords: NF-kappa B (NF-KB); Pox Viruses; Signal Transduction; TRAF6; Viral Protein; p38 MAPK.
Figures
References
-
- DiPerna G., Stack J., Bowie A. G., Boyd A., Kotwal G., Zhang Z., Arvikar S., Latz E., Fitzgerald K. A., Marshall W. L. (2004) Poxvirus protein N1L targets the I-κB kinase complex, inhibits signaling to NF-κB by the tumor necrosis factor superfamily of receptors, and inhibits NF-κB and IRF3 signaling by toll-like receptors. J. Biol. Chem. 279, 36570–36578 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
