

Lysophosphatidic acid (LPA) and its receptor, LPA1 , influence embryonic schwann cell migration, myelination, and cell-to-axon segregation
- PMID: 24115248
- PMCID: PMC3941654
- DOI: 10.1002/glia.22572
Lysophosphatidic acid (LPA) and its receptor, LPA1 , influence embryonic schwann cell migration, myelination, and cell-to-axon segregation
Abstract
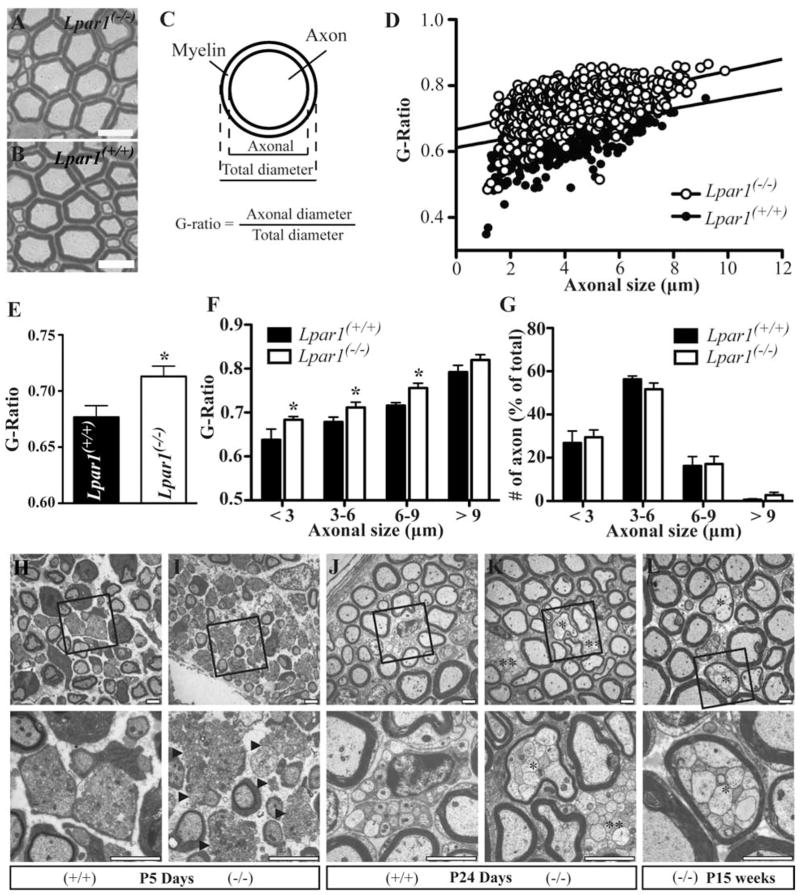
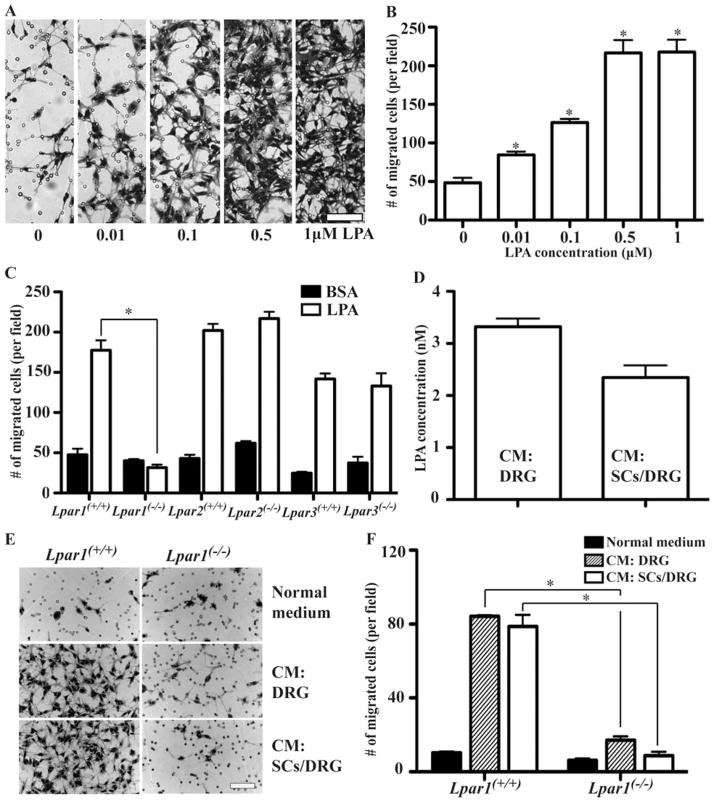
Schwann cell (SC) migration is an important step preceding myelination and remyelination in the peripheral nervous system, and can be promoted by peptide factors like neuregulins. Here we present evidence that a lipid factor, lysophosphatidic acid (LPA), influences both SC migration and peripheral myelination through its cognate G protein-coupled receptor (GPCR) known as LPA1 . Ultrastructural analyses of peripheral nerves in mouse null-mutants for LPA1 showed delayed SC-to-axon segregation, polyaxonal myelination by single SCs, and thinner myelin sheaths. In primary cultures, LPA promoted SC migration through LPA1 , while analysis of conditioned media from purified dorsal root ganglia neurons using HPLC/MS supported the production of LPA by these neurons. The heterotrimeric G-alpha protein, Gαi , and the small GTPase, Rac1, were identified as important downstream signaling components of LPA1 . These results identify receptor mediated LPA signaling between neurons and SCs that promote SC migration and contribute to the normal development of peripheral nerves through effects on SC-axon segregation and myelination.
Keywords: Gi; LPA; Rac1; Schwann cell; myelination.
Copyright © 2013 Wiley Periodicals, Inc.
Figures




References
-
- Allard J, Barron S, Diaz J, Lubetzki C, Zalc B, Schwartz JC, Sokoloff P. A rat G protein-coupled receptor selectively expressed in myelin-forming cells. Eur J Neurosci. 1998;10:1045–1053. - PubMed
-
- Anliker B, Chun J. Lysophospholipid G protein-coupled receptors. J Biol Chem. 2004;279:20555–20558. - PubMed
-
- Benard V, Bokoch GM. Assay of Cdc42, Rac, and Rho GTPase activation by affinity methods. Methods Enzymol. 2002;345:349–359. - PubMed
-
- Benninger Y, Thurnherr T, Pereira JA, Krause S, Wu X, Chrostek-Grashoff A, Herzog D, Nave KA, Franklin RJ, Meijer D, et al. Essential and distinct roles for cdc42 and rac1 in the regulation of Schwann cell biology during AQ2 peripheral nervous system development. J Cell Biol. 2007;177(6):1051–61. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
