

Fueling immunity: insights into metabolism and lymphocyte function
- PMID: 24115444
- PMCID: PMC4486656
- DOI: 10.1126/science.1242454
Fueling immunity: insights into metabolism and lymphocyte function
Abstract
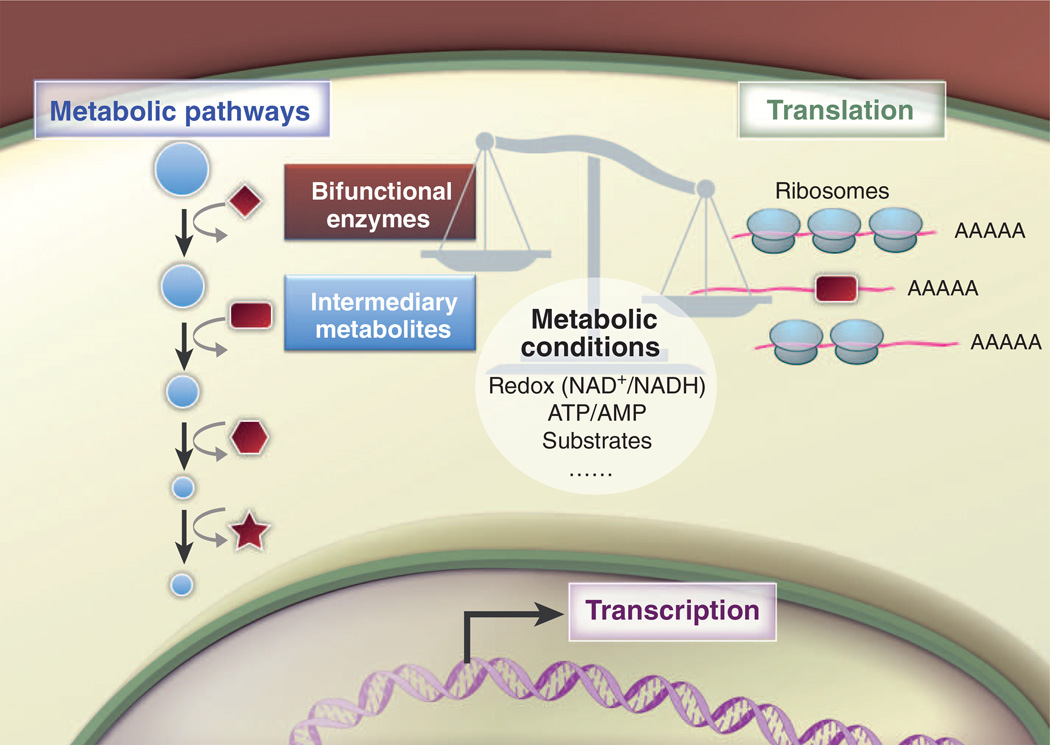
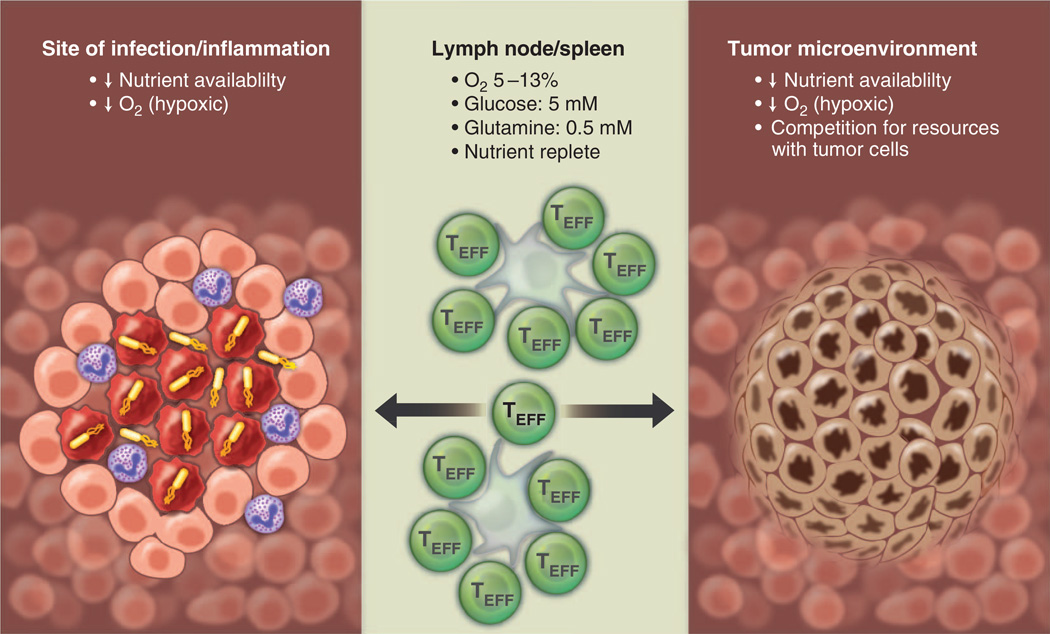
Lymphocytes face major metabolic challenges upon activation. They must meet the bioenergetic and biosynthetic demands of increased cell proliferation and also adapt to changing environmental conditions, in which nutrients and oxygen may be limiting. An emerging theme in immunology is that metabolic reprogramming and lymphocyte activation are intricately linked. However, why T cells adopt specific metabolic programs and the impact that these programs have on T cell function and, ultimately, immunological outcome remain unclear. Research on tumor cell metabolism has provided valuable insight into metabolic pathways important for cell proliferation and the influence of metabolites themselves on signal transduction and epigenetic programming. In this Review, we highlight emerging concepts regarding metabolic reprogramming in proliferating cells and discuss their potential impact on T cell fate and function.
Figures
References
-
- Wang R, Green DR. Metabolic checkpoints in activated T cells. Nat. Immunol. 2012;13:907–915. pmid: 22990888. - PubMed
-
- O’Neill LA, Hardie DG. Metabolism of inflammation limited by AMPK and pseudo-starvation. Nature. 2013;493:346–355. pmid: 23325217. - PubMed
-
- Greiner EF, Guppy M, Brand K. Glucose is essential for proliferation and the glycolytic enzyme induction that provokes a transition to glycolytic energy production. J. Biol. Chem. 1994;269:31484–31490. pmid: 7989314. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
