

Regulation of T-cell receptor signaling by the actin cytoskeleton and poroelastic cytoplasm
- PMID: 24117819
- PMCID: PMC3831008
- DOI: 10.1111/imr.12120
Regulation of T-cell receptor signaling by the actin cytoskeleton and poroelastic cytoplasm
Abstract
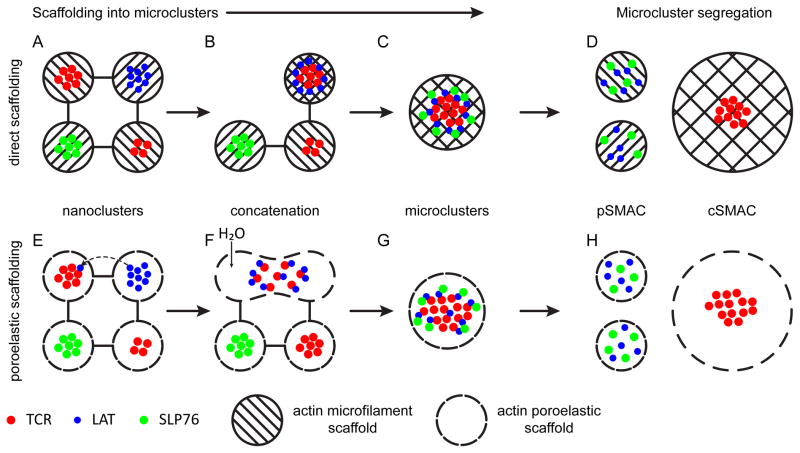
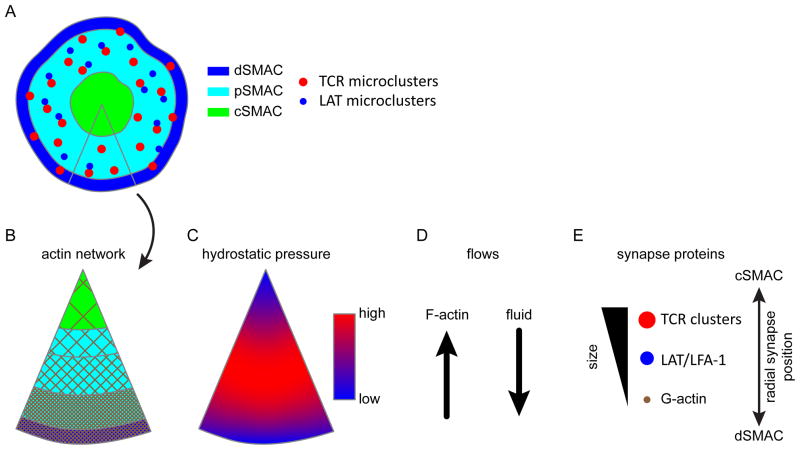
The actin cytoskeleton plays essential roles in modulating T-cell activation. Most models of T-cell receptor (TCR) triggering signalosome assembly and immune synapse formation invoke actin-dependent mechanisms. As T cells are constitutively motile cells, TCR triggering and signaling occur against a cytoskeletal backdrop that is constantly remodeling. While the interplay between actin dynamics and TCR signaling have been the focus of research for many years, much of the work in T cells has considered actin largely for its 'scaffolding' function. We examine the roles of the actin cytoskeleton in TCR signaling and immune synapse formation with an emphasis on how poroelasticity, an ensemble feature of actin dynamics with the cytosol, relates to how T cells respond to stimulation.
Keywords: T cells; T-cell receptors; signal transduction.
© 2013 John Wiley & Sons A/S. Published by John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare that they have no financial conflict of interest.
Figures
References
-
- Valitutti S, Muller S, Cella M, Padovan E, Lanzavecchia A. Serial triggering of many T-cell receptors by a few peptide–MHC complexes. Nature. 1995;375:148–151. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
