

Structure of the CRISPR interference complex CSM reveals key similarities with cascade
- PMID: 24119402
- PMCID: PMC3807668
- DOI: 10.1016/j.molcel.2013.08.020
Structure of the CRISPR interference complex CSM reveals key similarities with cascade
Abstract
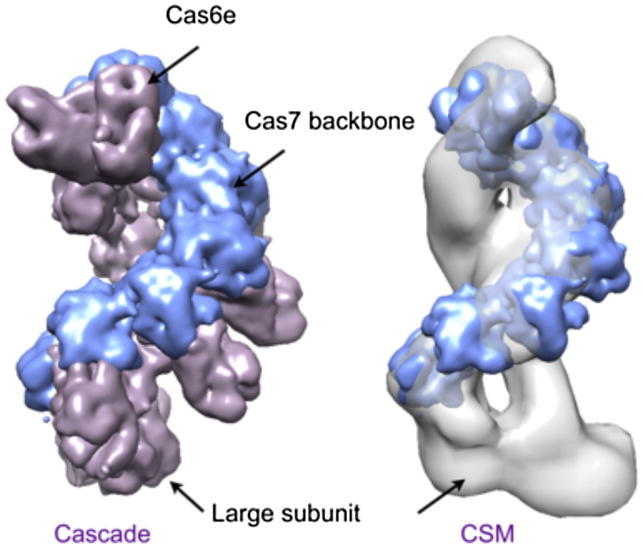
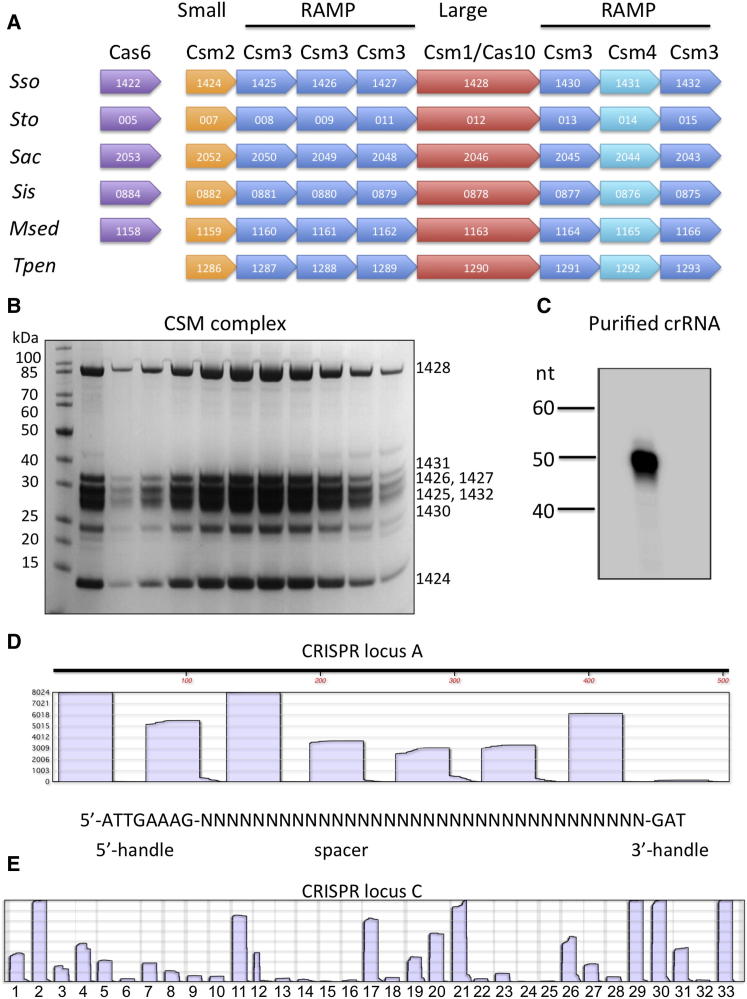
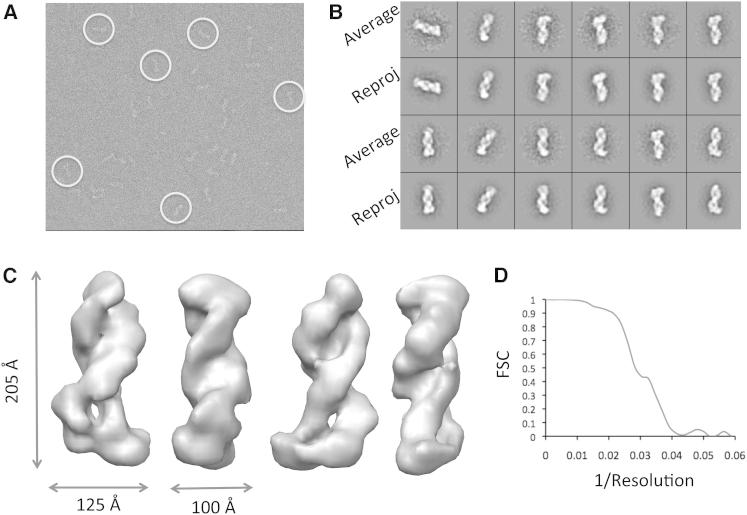
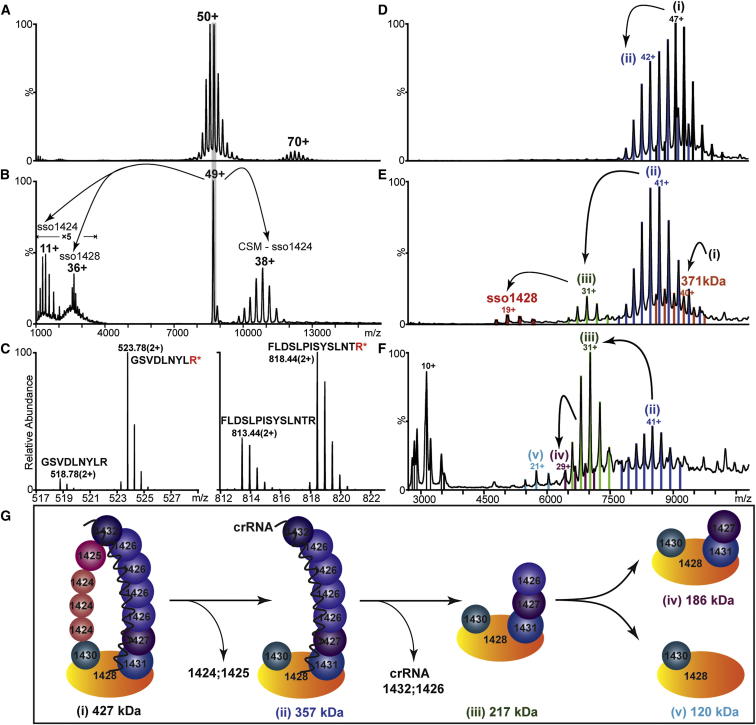
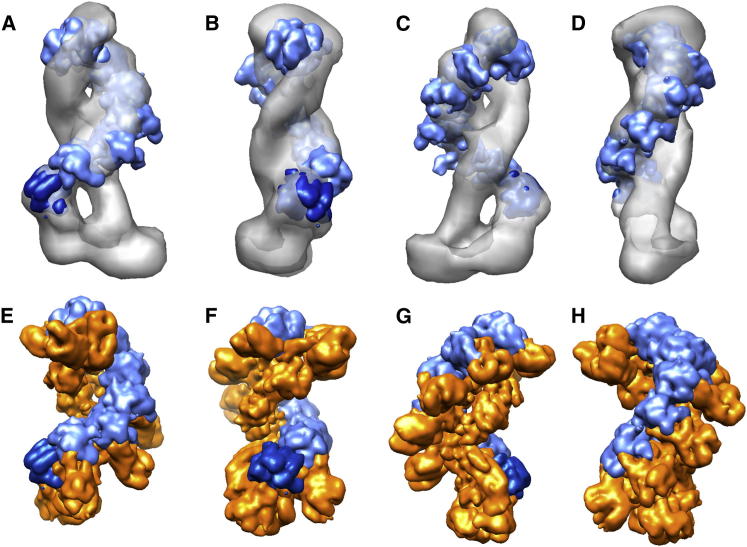
The Clustered Regularly Interspaced Palindromic Repeats (CRISPR) system is an adaptive immune system in prokaryotes. Interference complexes encoded by CRISPR-associated (cas) genes utilize small RNAs for homology-directed detection and subsequent degradation of invading genetic elements, and they have been classified into three main types (I-III). Type III complexes share the Cas10 subunit but are subclassifed as type IIIA (CSM) and type IIIB (CMR), depending on their specificity for DNA or RNA targets, respectively. The role of CSM in limiting the spread of conjugative plasmids in Staphylococcus epidermidis was first described in 2008. Here, we report a detailed investigation of the composition and structure of the CSM complex from the archaeon Sulfolobus solfataricus, using a combination of electron microscopy, mass spectrometry, and deep sequencing. This reveals a three-dimensional model for the CSM complex that includes a helical component strikingly reminiscent of the backbone structure of the type I (Cascade) family.
Copyright © 2013 The Authors. Published by Elsevier Inc. All rights reserved.
Figures

Comment in
-
Same same but different: new structural insight into CRISPR-Cas complexes.Mol Cell. 2013 Oct 10;52(1):4-7. doi: 10.1016/j.molcel.2013.09.023. Mol Cell. 2013. PMID: 24119398
References
-
- Alber F., Kim M.F., Sali A. Structural characterization of assemblies from overall shape and subcomplex compositions. Structure. 2005;13:435–445. - PubMed
-
- Esser D., Pham T.K., Reimann J., Albers S.V., Siebers B., Wright P.C. Change of carbon source causes dramatic effects in the phospho-proteome of the archaeon Sulfolobus solfataricus. J. Proteome Res. 2012;11:4823–4833. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- 087658/Wellcome Trust/United Kingdom
- BB/K000314/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/J005665/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/J005673/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- WT087658MA/WT_/Wellcome Trust/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
