

Exercise induces hippocampal BDNF through a PGC-1α/FNDC5 pathway
- PMID: 24120943
- PMCID: PMC3980968
- DOI: 10.1016/j.cmet.2013.09.008
Exercise induces hippocampal BDNF through a PGC-1α/FNDC5 pathway
Abstract
Exercise can improve cognitive function and has been linked to the increased expression of brain-derived neurotrophic factor (BDNF). However, the underlying molecular mechanisms driving the elevation of this neurotrophin remain unknown. Here we show that FNDC5, a previously identified muscle protein that is induced in exercise and is cleaved and secreted as irisin, is also elevated by endurance exercise in the hippocampus of mice. Neuronal Fndc5 gene expression is regulated by PGC-1α, and Pgc1a(-/-) mice show reduced Fndc5 expression in the brain. Forced expression of FNDC5 in primary cortical neurons increases Bdnf expression, whereas RNAi-mediated knockdown of FNDC5 reduces Bdnf. Importantly, peripheral delivery of FNDC5 to the liver via adenoviral vectors, resulting in elevated blood irisin, induces expression of Bdnf and other neuroprotective genes in the hippocampus. Taken together, our findings link endurance exercise and the important metabolic mediators, PGC-1α and FNDC5, with BDNF expression in the brain.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
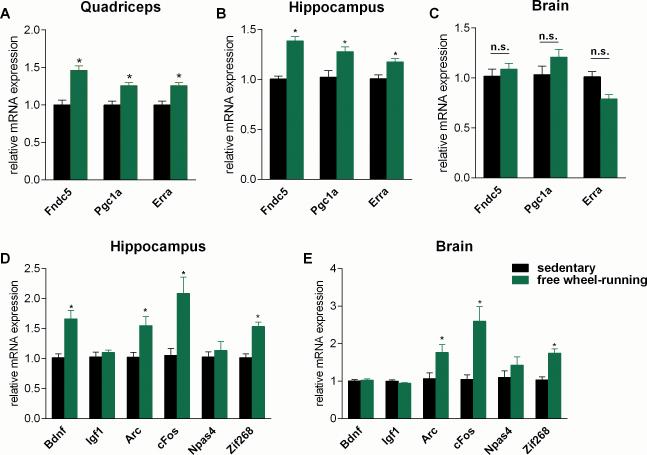
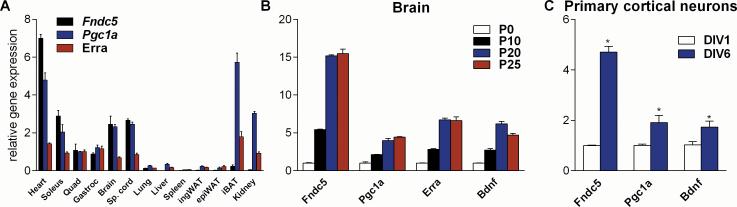
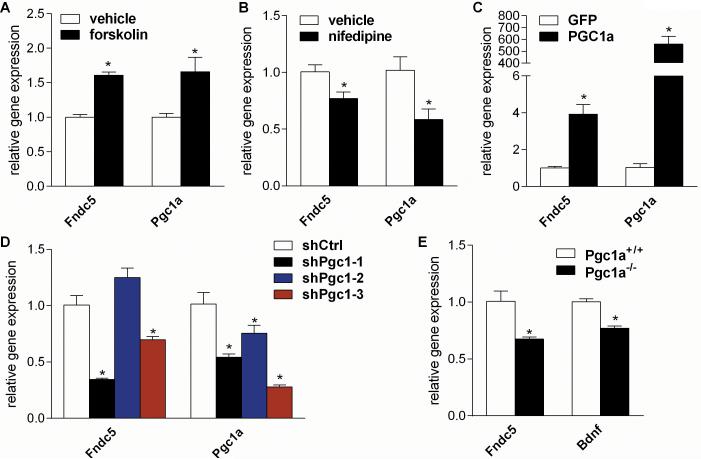
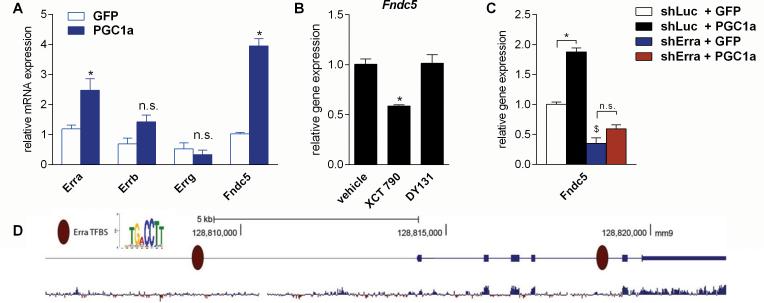
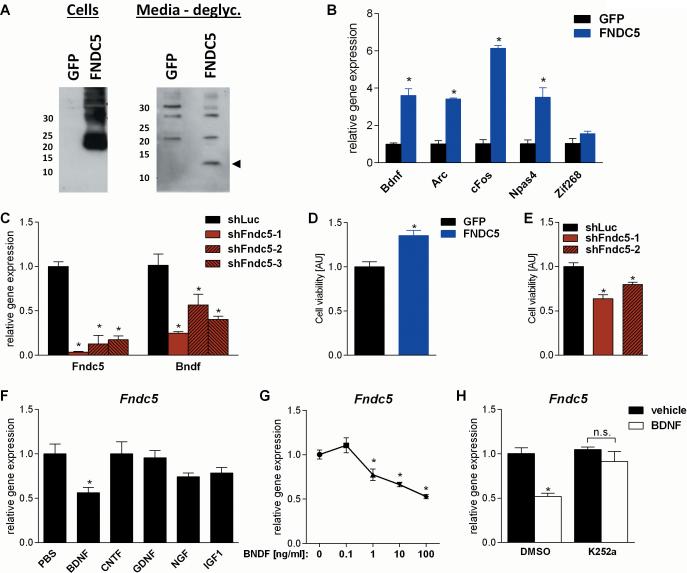
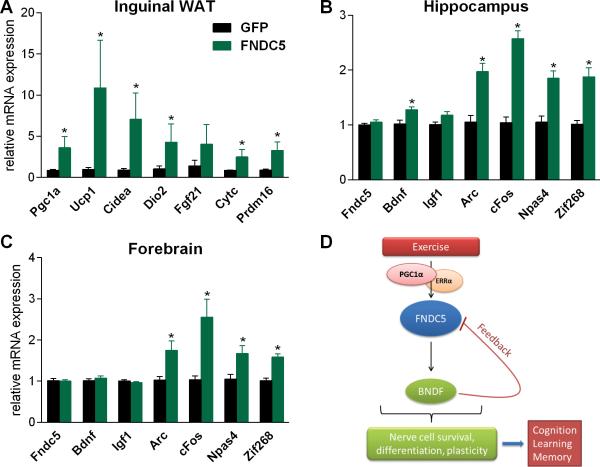
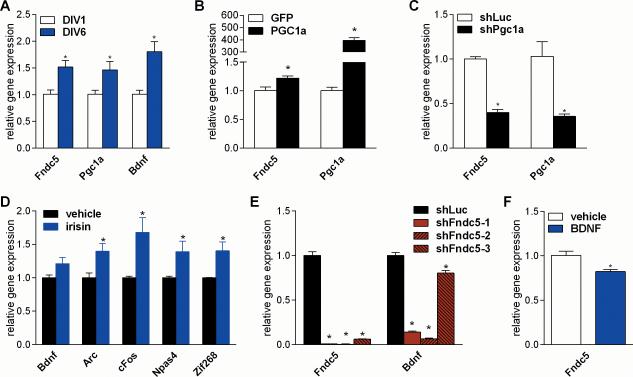
Comment in
-
BDNF (I)rising from exercise.Cell Metab. 2013 Nov 5;18(5):612-4. doi: 10.1016/j.cmet.2013.10.008. Cell Metab. 2013. PMID: 24206660 Free PMC article.
References
-
- Arany Z, Foo SY, Ma Y, Ruas JL, Bommi-Reddy A, Girnun G, Cooper M, Laznik D, Chinsomboon J, Rangwala SM, et al. HIF-independent regulation of VEGF and angiogenesis by the transcriptional coactivator PGC-1alpha. Nature. 2008;451:1008–1012. - PubMed
-
- Arida RM, Cavalheiro EA, da Silva AC, Scorza FA. Physical activity and epilepsy: proven and predicted benefits. Sports medicine (Auckland, NZ) 2008;38:607–615. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
