

Evolutionary origin of the mitochondrial cholesterol transport machinery reveals a universal mechanism of steroid hormone biosynthesis in animals
- PMID: 24124589
- PMCID: PMC3790746
- DOI: 10.1371/journal.pone.0076701
Evolutionary origin of the mitochondrial cholesterol transport machinery reveals a universal mechanism of steroid hormone biosynthesis in animals
Abstract
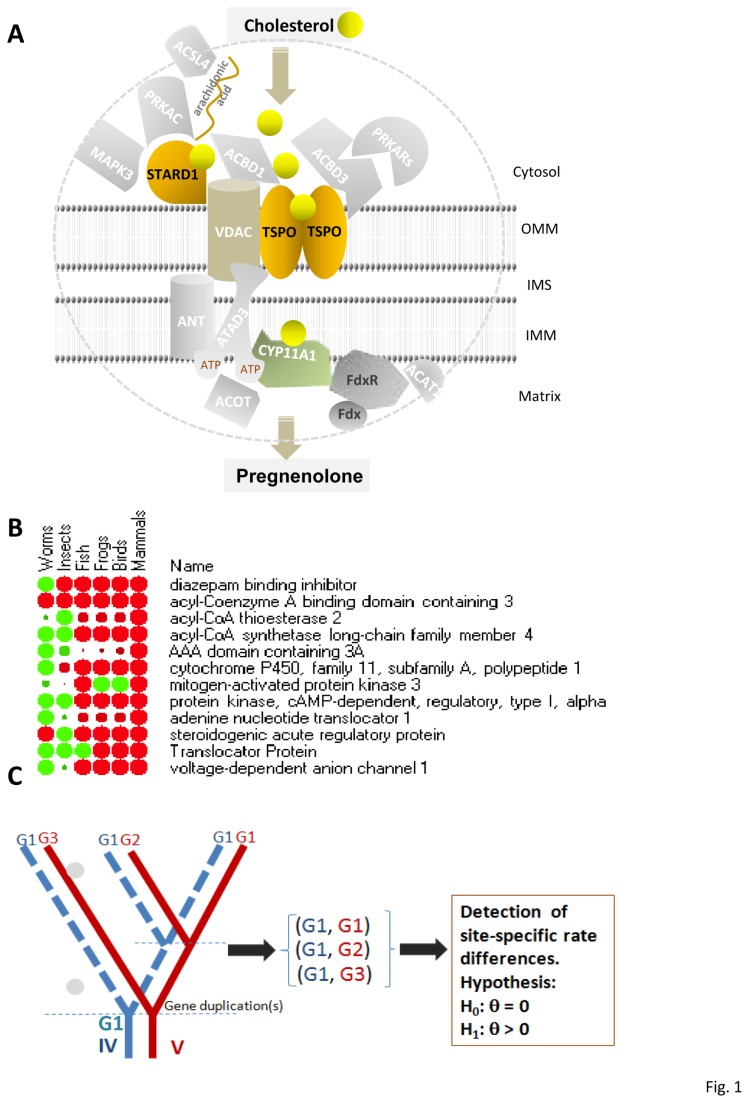
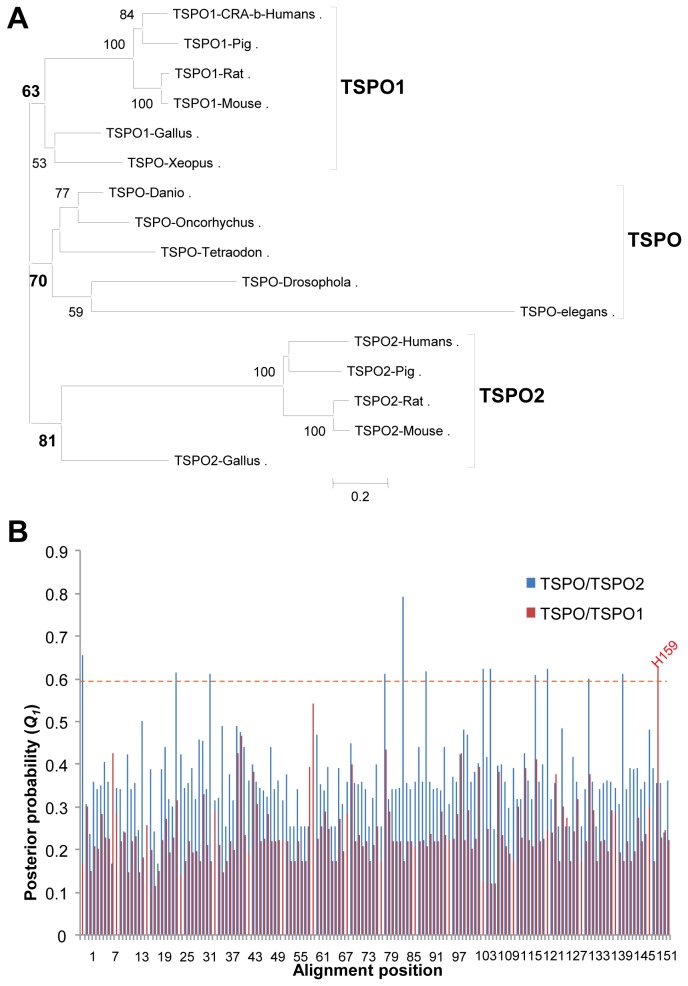
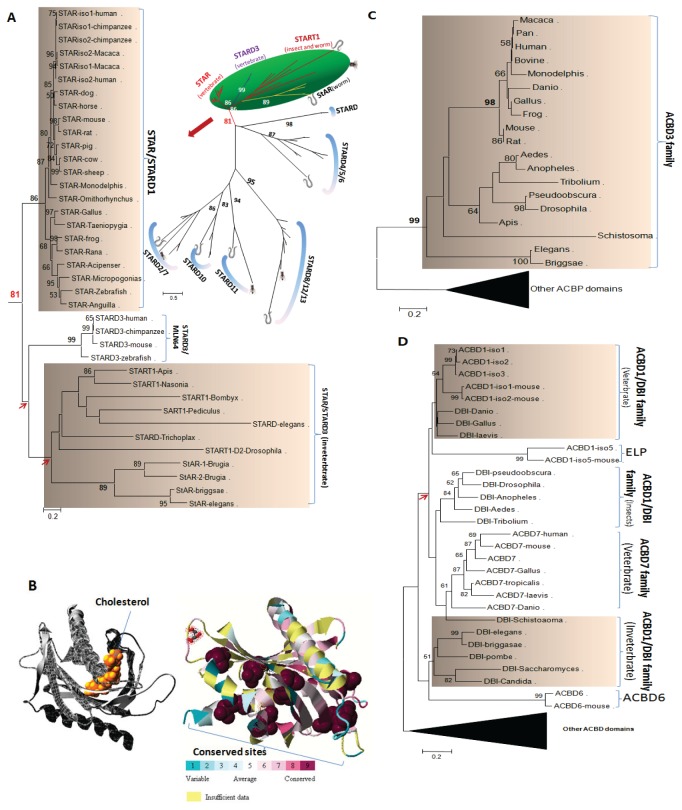
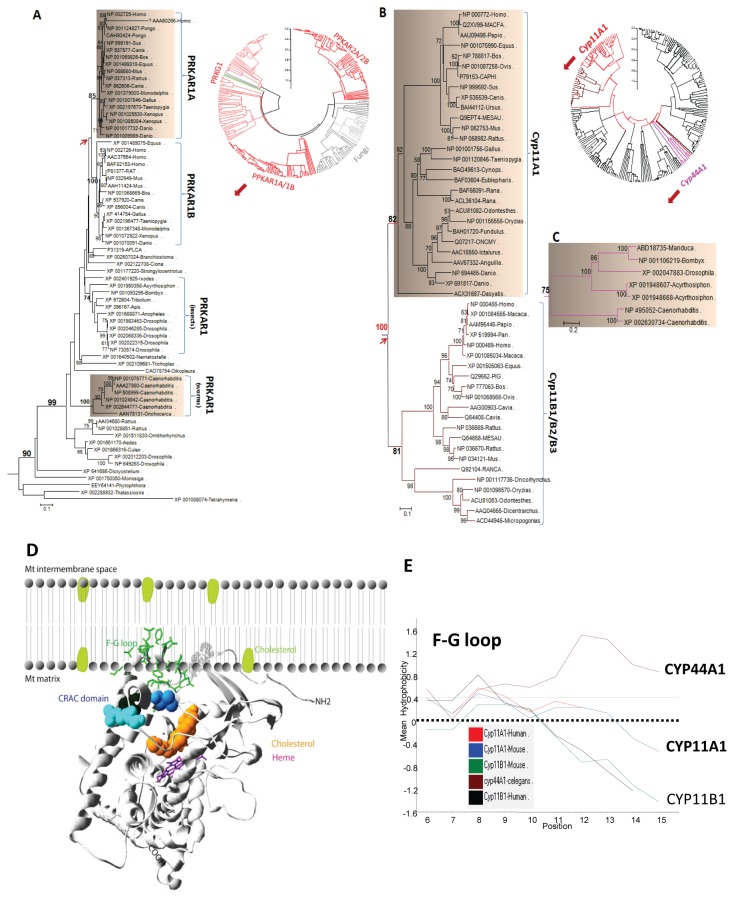
Steroidogenesis begins with the transport of cholesterol from intracellular stores into mitochondria via a series of protein-protein interactions involving cytosolic and mitochondrial proteins located at both the outer and inner mitochondrial membranes. In adrenal glands and gonads, this process is accelerated by hormones, leading to the production of high levels of steroids that control tissue development and function. A hormone-induced multiprotein complex, the transduceosome, was recently identified, and is composed of cytosolic and outer mitochondrial membrane proteins that control the rate of cholesterol entry into the outer mitochondrial membrane. More recent studies unveiled the steroidogenic metabolon, a bioactive, multimeric protein complex that spans the outer-inner mitochondrial membranes and is responsible for hormone-induced import, segregation, targeting, and metabolism of cholesterol by cytochrome P450 family 11 subfamily A polypeptide 1 (CYP11A1) in the inner mitochondrial membrane. The availability of genome information allowed us to systematically explore the evolutionary origin of the proteins involved in the mitochondrial cholesterol transport machinery (transduceosome, steroidogenic metabolon, and signaling proteins), trace the original archetype, and predict their biological functions by molecular phylogenetic and functional divergence analyses, protein homology modeling and molecular docking. Although most members of these complexes have a history of gene duplication and functional divergence during evolution, phylogenomic analysis revealed that all vertebrates have the same functional complex members, suggesting a common mechanism in the first step of steroidogenesis. An archetype of the complex was found in invertebrates. The data presented herein suggest that the cholesterol transport machinery is responsible for steroidogenesis among all vertebrates and is evolutionarily conserved throughout the entire animal kingdom.
Conflict of interest statement
Figures
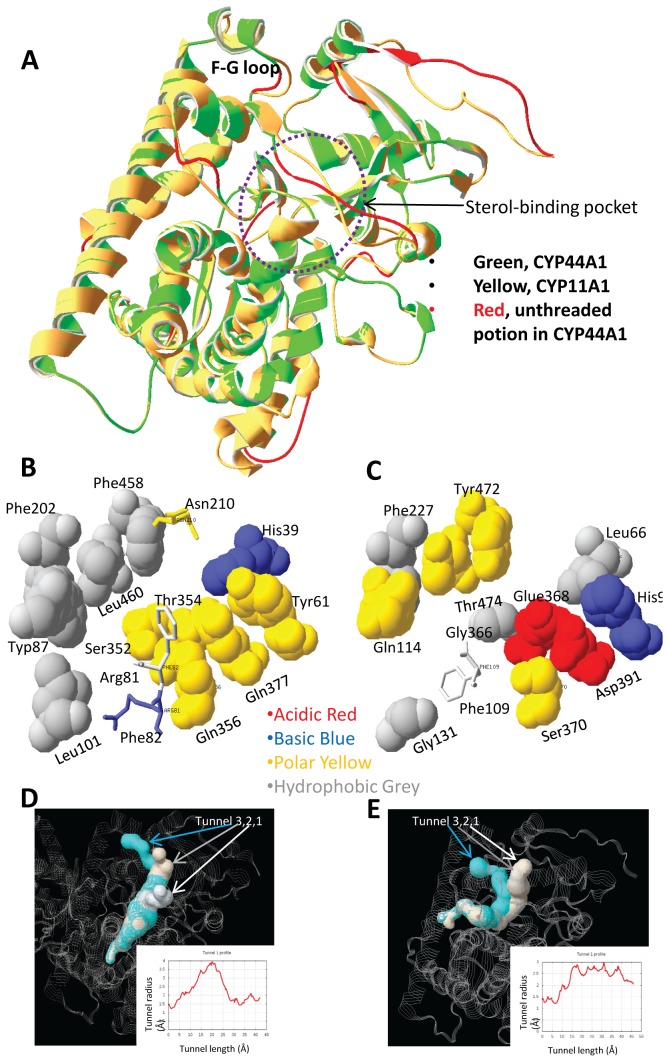
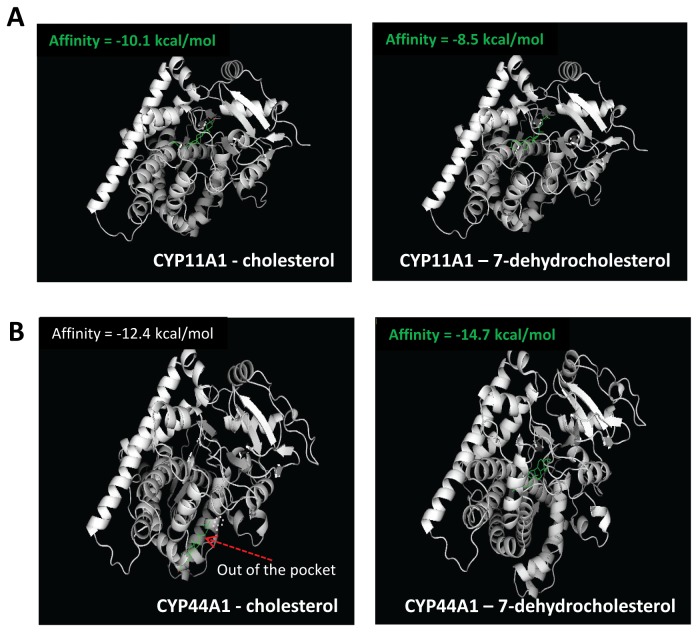

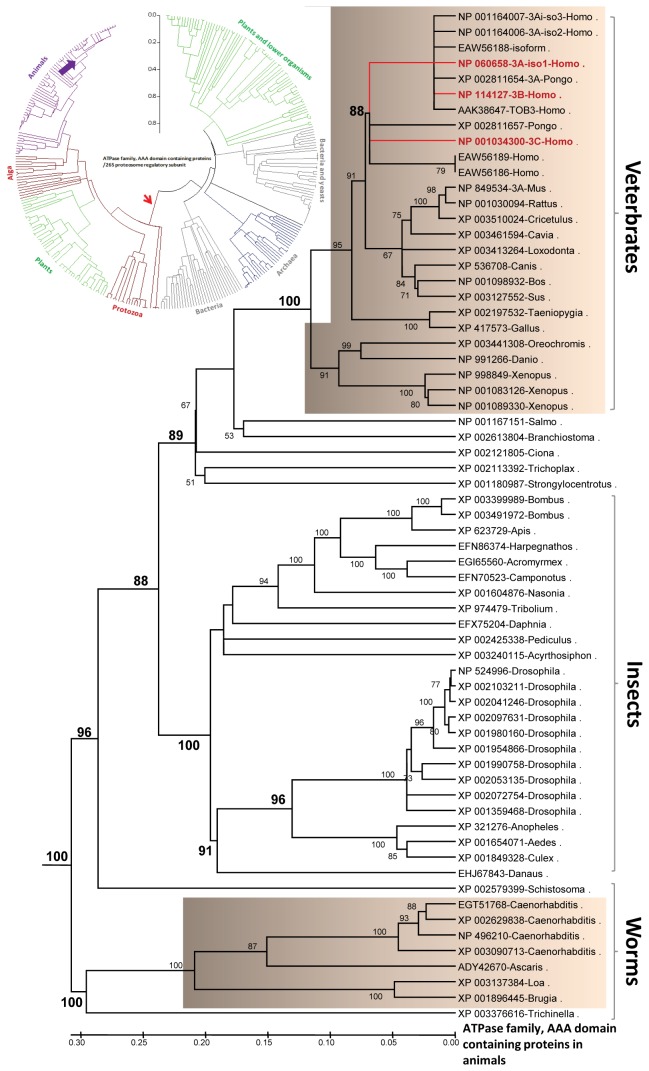
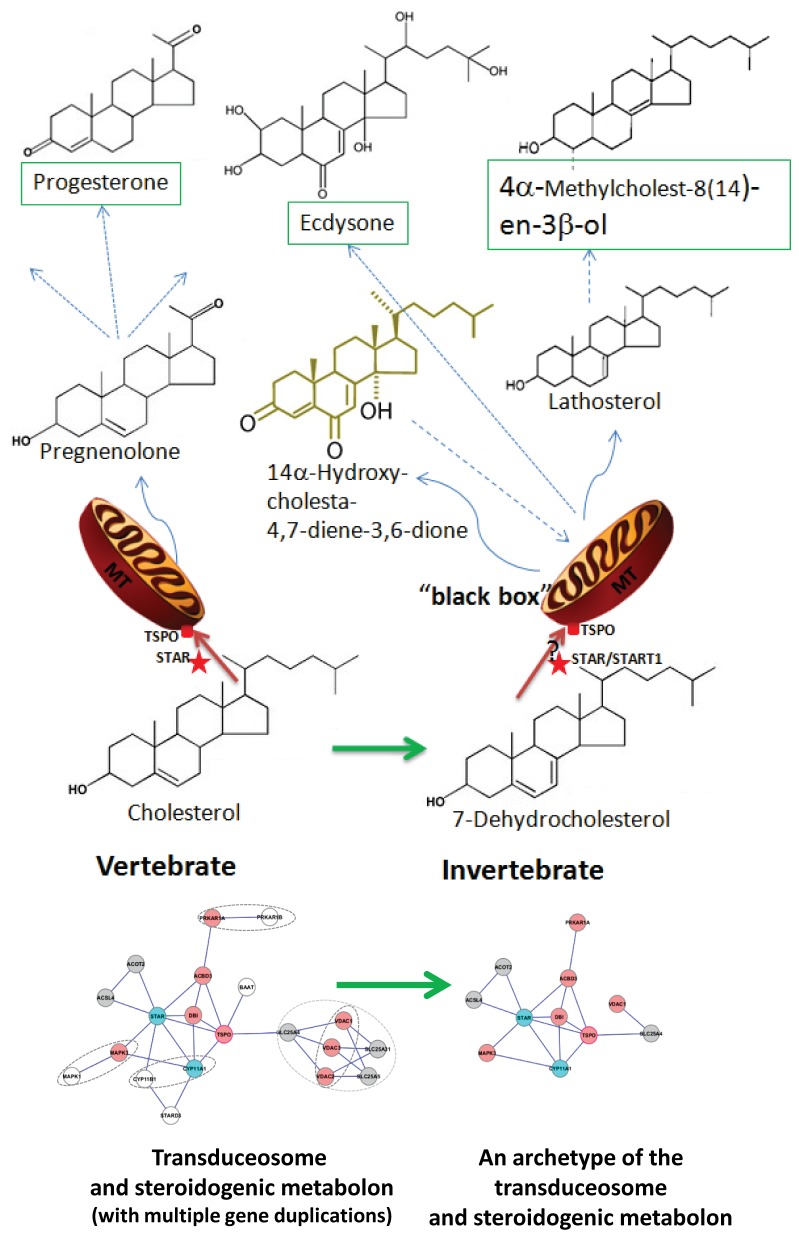
References
-
- Papadopoulos V, Baraldi M, Guilarte TR, Knudsen TB, Lacapère JJ et al. (2006) Translocator protein (18kDa): new nomenclature for the peripheral-type benzodiazepine receptor based on its structure and molecular function. Trends Pharmacol Sci 27: 402-409. doi: 10.1016/j.tips.2006.06.005. PubMed: 16822554. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
