

Couples, pairs, and clusters: mechanisms and implications of centromere associations in meiosis
- PMID: 24126501
- PMCID: PMC3969401
- DOI: 10.1007/s00412-013-0439-4
Couples, pairs, and clusters: mechanisms and implications of centromere associations in meiosis
Abstract
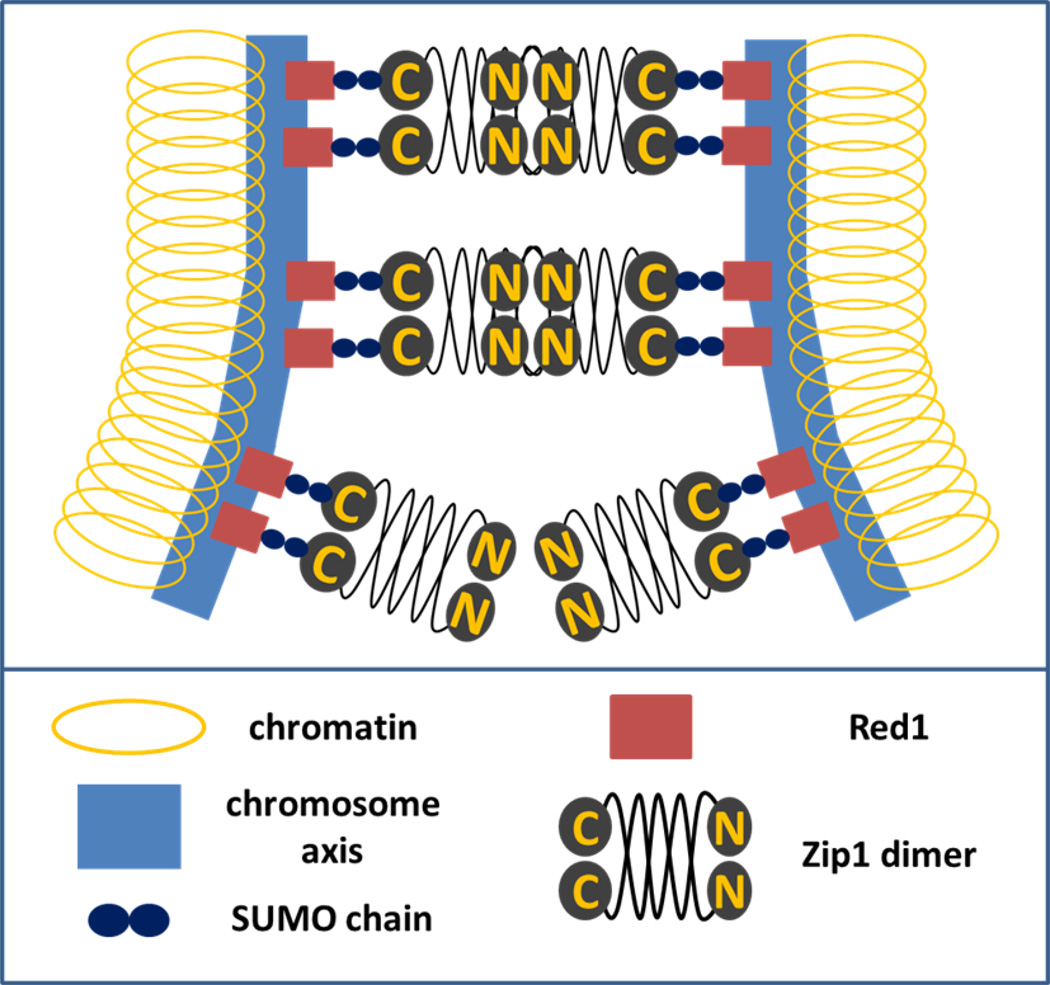
Observations of a wide range of organisms show that the centromeres form associations of pairs or small groups at different stages of meiotic prophase. Little is known about the functions or mechanisms of these associations, but in many cases, synaptonemal complex elements seem to play a fundamental role. Two main associations are observed: homology-independent associations very early in the meiotic program-sometimes referred to as centromere coupling-and a later association of homologous centromeres, referred to as centromere pairing or tethering. The later centromere pairing initiates during synaptonemal complex assembly, then persists after the dissolution of the synaptonemal complex. While the function of the homology-independent centromere coupling remains a mystery, centromere pairing appears to have a direct impact on the chromosome segregation fidelity of achiasmatic chromosomes. Recent work in yeast, Drosophila, and mice suggest that centromere pairing is a previously unappreciated, general meiotic feature that may promote meiotic segregation fidelity of the exchange and non-exchange chromosomes.
Figures




References
-
- Agarwal S, Roeder GS. Zip3 provides a link between recombination enzymes and synaptonemal complex proteins. Cell. 2000;102:245–255. - PubMed
-
- Bennett MD. Centromere arrangements in. Triticum aestivum and their relationship to synapsis. Heredity. 1979;43:157.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
