

Anti-CCR4 mAb selectively depletes effector-type FoxP3+CD4+ regulatory T cells, evoking antitumor immune responses in humans
- PMID: 24127572
- PMCID: PMC3816454
- DOI: 10.1073/pnas.1316796110
Anti-CCR4 mAb selectively depletes effector-type FoxP3+CD4+ regulatory T cells, evoking antitumor immune responses in humans
Abstract
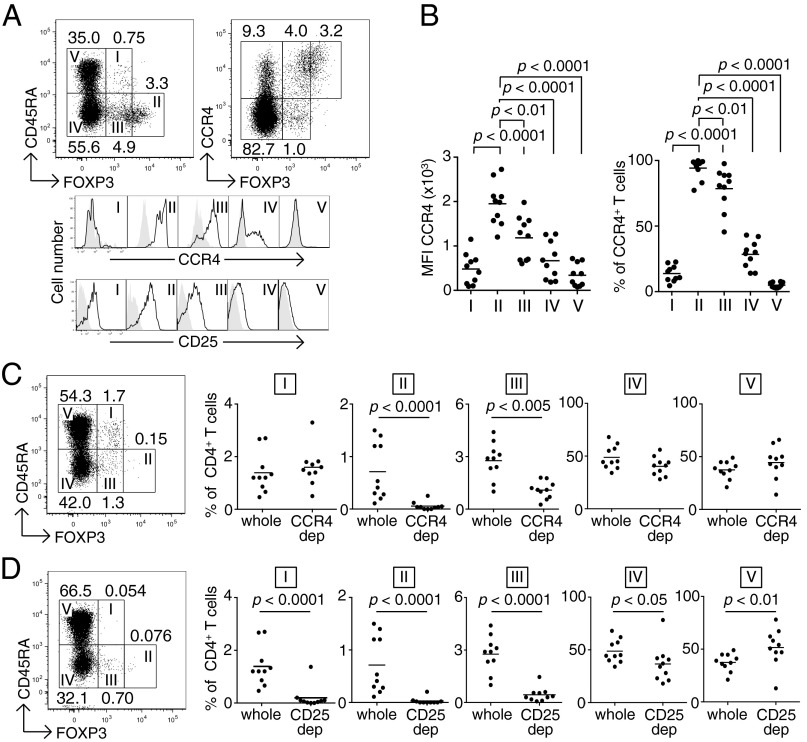
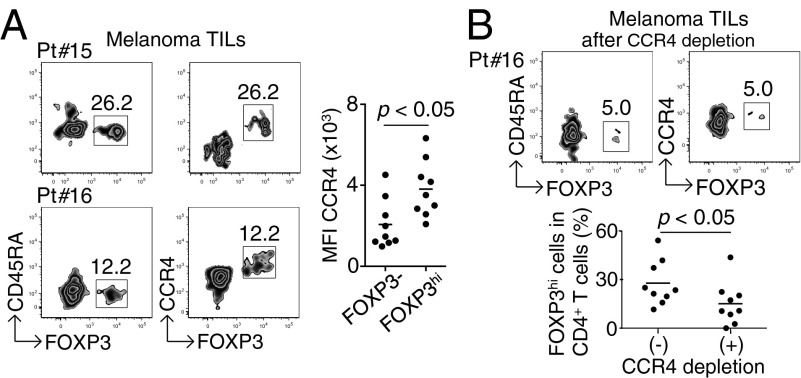
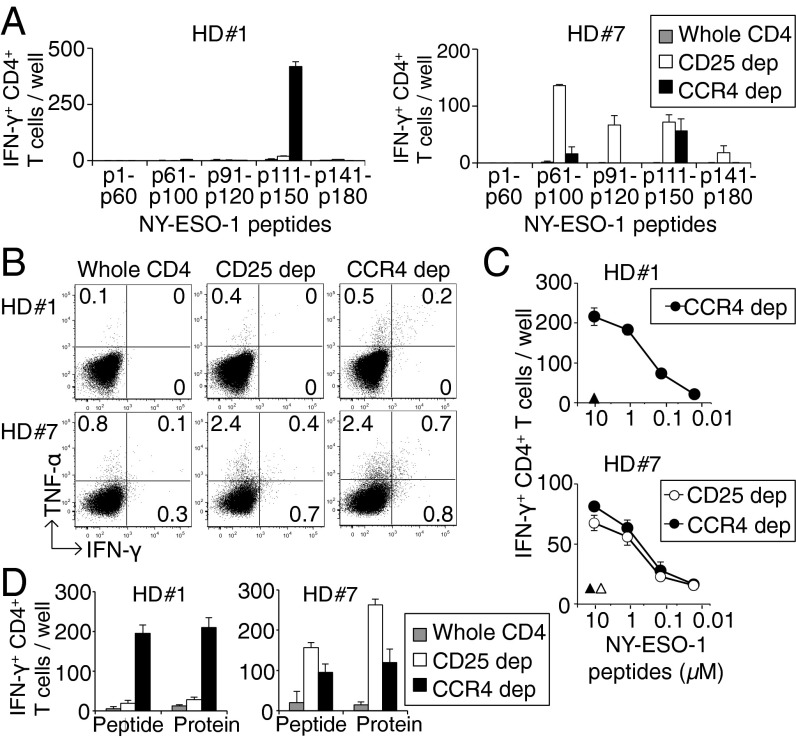
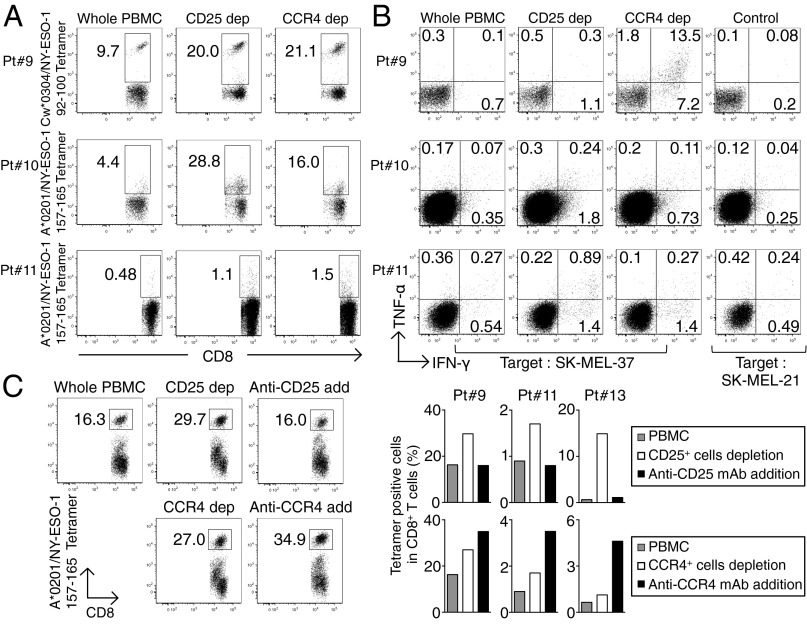
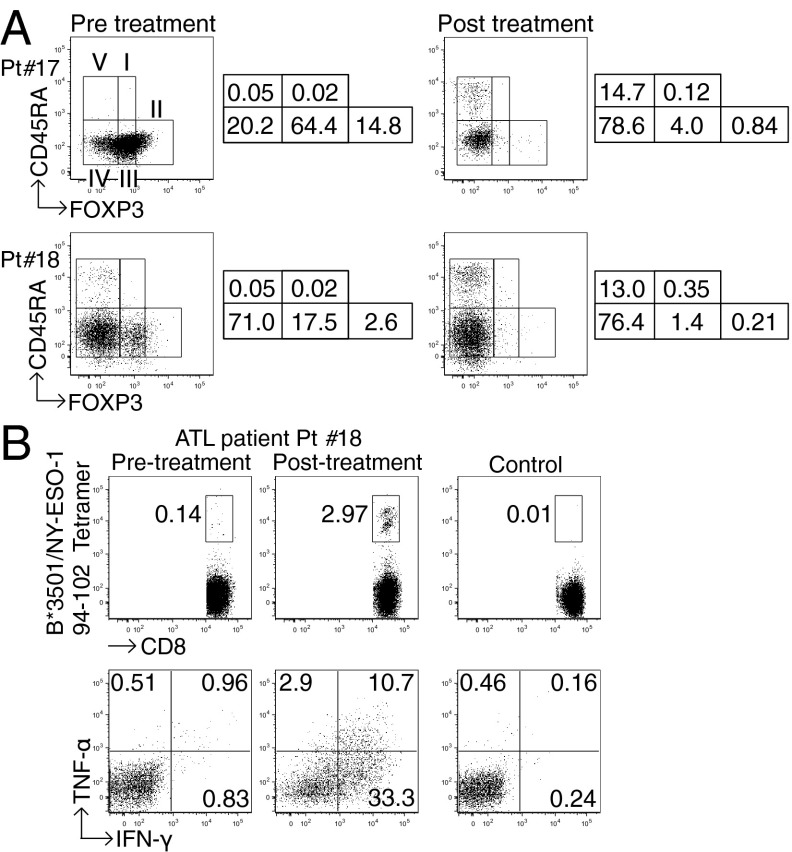
CD4(+) Treg cells expressing the transcription factor FOXP3 (forkhead box P3) are abundant in tumor tissues and appear to hinder the induction of effective antitumor immunity. A substantial number of T cells, including Treg cells, in tumor tissues and peripheral blood express C-C chemokine receptor 4 (CCR4). Here we show that CCR4 was specifically expressed by a subset of terminally differentiated and most suppressive CD45RA(-)FOXP3(hi)CD4(+) Treg cells [designated effector Treg (eTreg) cells], but not by CD45RA(+)FOXP3(lo)CD4(+) naive Treg cells, in peripheral blood of healthy individuals and cancer patients. In melanoma tissues, CCR4(+) eTreg cells were predominant among tumor-infiltrating FOXP3(+) T cells and much higher in frequency compared with those in peripheral blood. With peripheral blood lymphocytes from healthy individuals and melanoma patients, ex vivo depletion of CCR4(+) T cells and subsequent in vitro stimulation of the depleted cell population with the cancer/testis antigen NY-ESO-1 efficiently induced NY-ESO-1-specific CD4(+) T cells. Nondepletion failed in the induction. The magnitude of the responses was comparable with total removal of FOXP3(+) Treg cells by CD25(+) T-cell depletion. CCR4(+) T-cell depletion also augmented in vitro induction of NY-ESO-1-specific CD8(+) T cells in melanoma patients. Furthermore, in vivo administration of anti-CCR4 mAb markedly reduced the eTreg-cell fraction and augmented NY-ESO-1-specific CD8(+) T-cell responses in an adult T-cell leukemia-lymphoma patient whose leukemic cells expressed NY-ESO-1. Collectively, these findings indicate that anti-CCR4 mAb treatment is instrumental for evoking and augmenting antitumor immunity in cancer patients by selectively depleting eTreg cells.
Keywords: cancer immunotherapy; immunomodulation.
Conflict of interest statement
Conflict of interest statement: H.N. received a research grant from Kyowa Hakko Kirin Co., Ltd.
Figures
Comment in
-
Tumour immunology: dealing with regulators.Nat Rev Immunol. 2013 Dec;13(12):846-7. doi: 10.1038/nri3569. Epub 2013 Nov 15. Nat Rev Immunol. 2013. PMID: 24232463 No abstract available.
References
-
- Sakaguchi S. Naturally arising CD4+ regulatory t cells for immunologic self-tolerance and negative control of immune responses. Annu Rev Immunol. 2004;22:531–562. - PubMed
-
- Sakaguchi S, Miyara M, Costantino CM, Hafler DA. FOXP3+ regulatory T cells in the human immune system. Nat Rev Immunol. 2010;10(7):490–500. - PubMed
-
- Kawakami Y, Rosenberg SA. Human tumor antigens recognized by T-cells. Immunol Res. 1997;16(4):313–339. - PubMed
-
- Scanlan MJ, Gure AO, Jungbluth AA, Old LJ, Chen YT. Cancer/testis antigens: An expanding family of targets for cancer immunotherapy. Immunol Rev. 2002;188:22–32. - PubMed
-
- Boon T, Coulie PG, Van den Eynde BJ, van der Bruggen P. Human T cell responses against melanoma. Annu Rev Immunol. 2006;24:175–208. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
