

Mutation in transforming growth factor beta induced protein associated with granular corneal dystrophy type 1 reduces the proteolytic susceptibility through local structural stabilization
- PMID: 24129074
- PMCID: PMC4162128
- DOI: 10.1016/j.bbapap.2013.10.008
Mutation in transforming growth factor beta induced protein associated with granular corneal dystrophy type 1 reduces the proteolytic susceptibility through local structural stabilization
Abstract
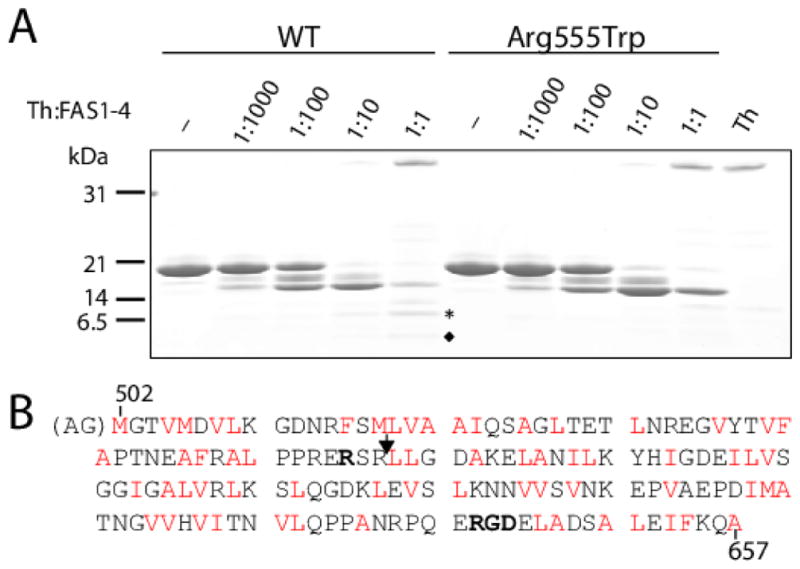
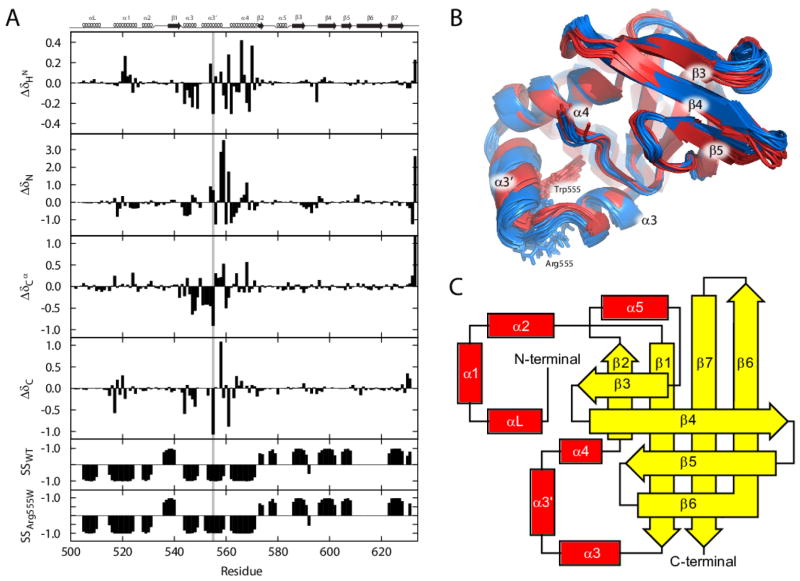
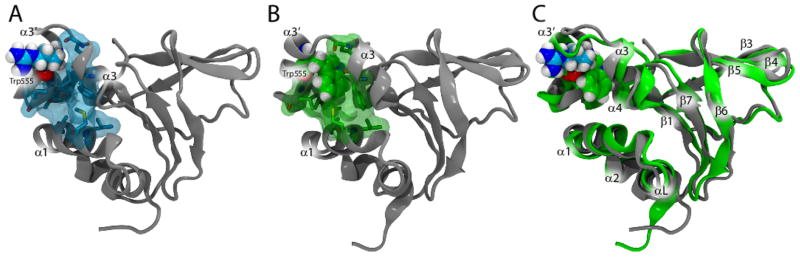
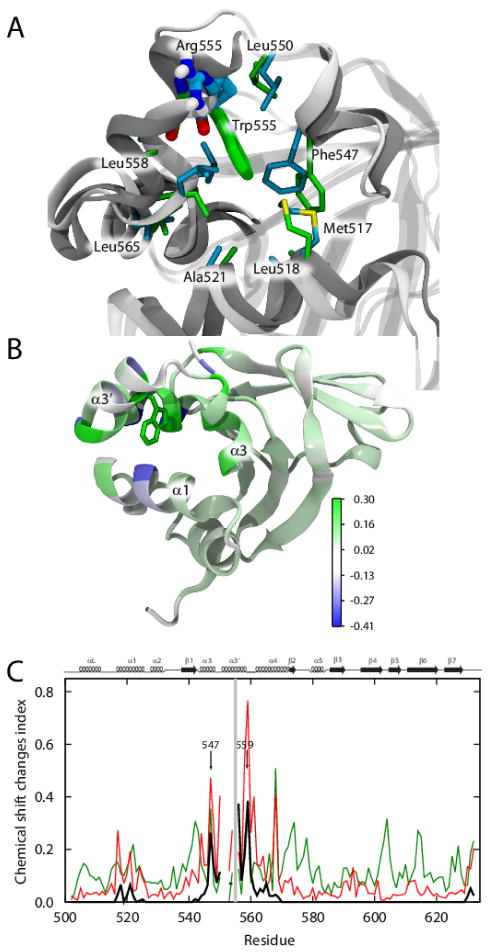
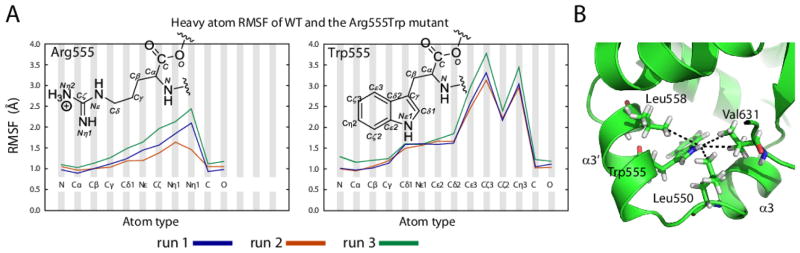
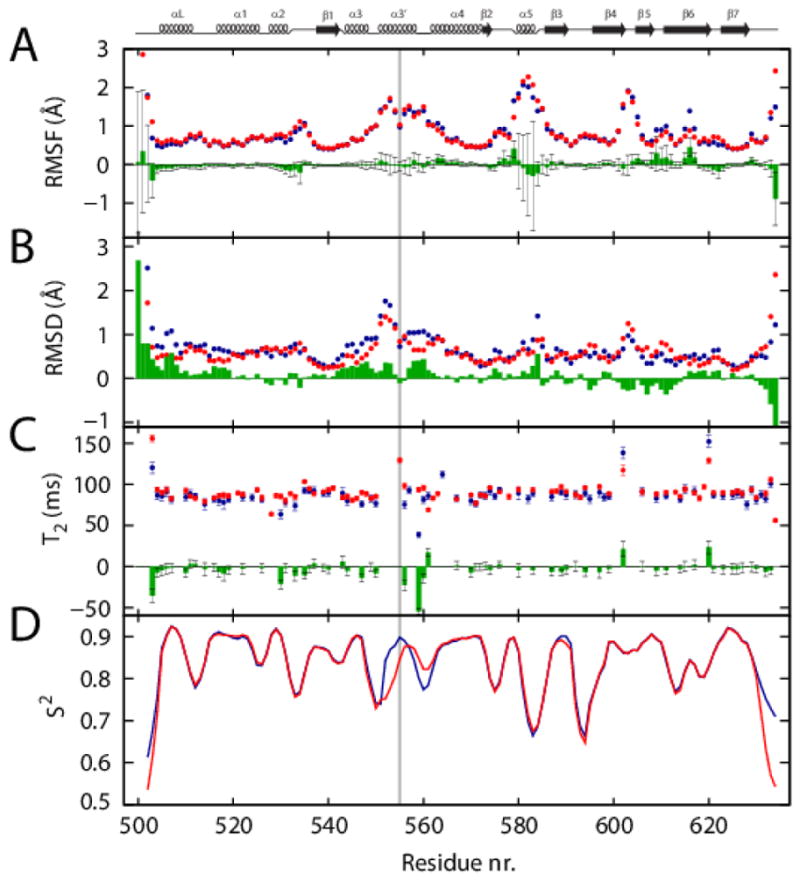
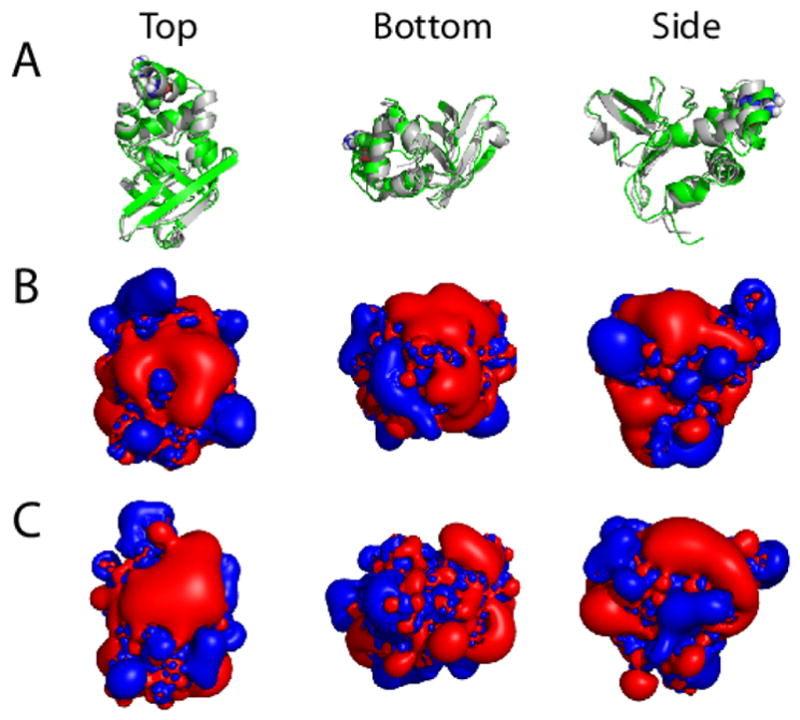
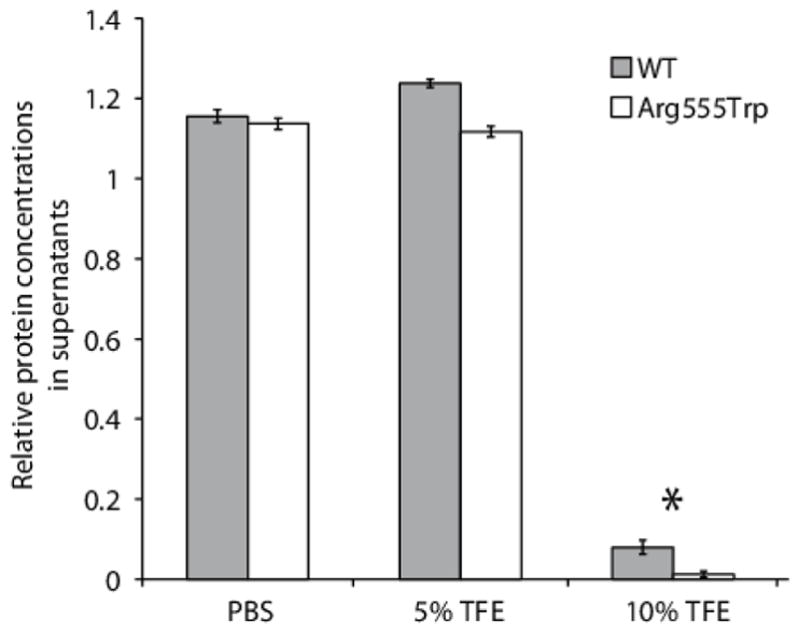
Hereditary mutations in the transforming growth factor beta induced (TGFBI) gene cause phenotypically distinct corneal dystrophies characterized by protein deposition in cornea. We show here that the Arg555Trp mutant of the fourth fasciclin 1 (FAS1-4) domain of the protein (TGFBIp/keratoepithelin/βig-h3), associated with granular corneal dystrophy type 1, is significantly less susceptible to proteolysis by thermolysin and trypsin than the WT domain. High-resolution liquid-state NMR of the WT and Arg555Trp mutant FAS1-4 domains revealed very similar structures except for the region around position 555. The Arg555Trp substitution causes Trp555 to be buried in an otherwise empty hydrophobic cavity of the FAS1-4 domain. The first thermolysin cleavage in the core of the FAS1-4 domain occurs on the N-terminal side of Leu558 adjacent to the Arg555 mutation. MD simulations indicated that the C-terminal end of helix α3' containing this cleavage site is less flexible in the mutant domain, explaining the observed proteolytic resistance. This structural change also alters the electrostatic properties, which may explain increased propensity of the mutant to aggregate in vitro with 2,2,2-trifluoroethanol. Based on our results we propose that the Arg555Trp mutation disrupts the normal degradation/turnover of corneal TGFBIp, leading to accumulation and increased propensity to aggregate through electrostatic interactions.
Keywords: 2,2-Dimethyl-2-silapentane-5-sulfonate; Corneal dystrophy; DSS; EMI; EMILIN-1 domain; FAS1; FAS1-4; GCD; IPTG; LB; LCD; Lysogeny broth; MD; NMR; NMR structure; OD; PDB; Protein Data Bank; Protein aggregation; Protein misfolding diseases; Proteolytic degradation; RDC; RMSD; RMSF; SUMO; TBCD; TGFBI; TGFBIp; Thiel–Behnke corneal dystrophy; Transforming growth factor beta induced protein (TGFBIp); WT; fasciclin 1 domain; fourth FAS1 domain of TGFBIp; granular corneal dystrophy; isopropyl β-d-1-thiogalactopyranoside; lattice corneal dystrophy; molecular dynamics; nuclear magnetic resonance; optical density; residual dipolar couplings; root-mean-square deviation; root-mean-square fluctuation; small ubiquitin-like modifier; transforming growth factor beta induced gene; transforming growth factor beta induced protein; wild-type.
© 2013.
Figures
References
-
- Kitahama S, Gibson MA, Hatzinikolas G, Hay S, Kuliwaba JL, Evdokiou A, Atkins GJ, Findlay DM. Expression of fibrillins and other microfibril-associated proteins in human bone and osteoblast-like cells. Bone. 2000;27:61–67. - PubMed
-
- LeBaron RG, Bezverkov KI, Zimber MP, Pavelec R, Skonier J, Purchio AF. Beta IG-H3, a novel secretory protein inducible by transforming growth factor-beta, is present in normal skin and promotes the adhesion and spreading of dermal fibroblasts in vitro. J Invest Dermatol. 1995;104:844–849. - PubMed
-
- Escribano J, Hernando N, Ghosh S, Crabb J, Coca-Prados M. cDNA from human ocular ciliary epithelium homologous to beta ig-h3 is preferentially expressed as an extracellular protein in the corneal epithelium. J Cell Physiol. 1994;160:511–521. - PubMed
-
- Ferguson JW, Thoma BS, Mikesh MF, Kramer RH, Bennett KL, Purchio A, Bellard BJ, LeBaron RG. The extracellular matrix protein betaIG-H3 is expressed at myotendinous junctions and supports muscle cell adhesion. Cell Tissue Res. 2003;313:93–105. - PubMed
-
- Doliana R, Bot S, Bonaldo P, Colombatti A. EMI, a novel cysteine-rich domain of EMILINs and other extracellular proteins, interacts with the gC1q domains and participates in multimerization. FEBS Letters. 2000;484:164–168. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
