

Factor H-dependent alternative pathway inhibition mediated by porin B contributes to virulence of Neisseria meningitidis
- PMID: 24129254
- PMCID: PMC3812710
- DOI: 10.1128/mBio.00339-13
Factor H-dependent alternative pathway inhibition mediated by porin B contributes to virulence of Neisseria meningitidis
Abstract
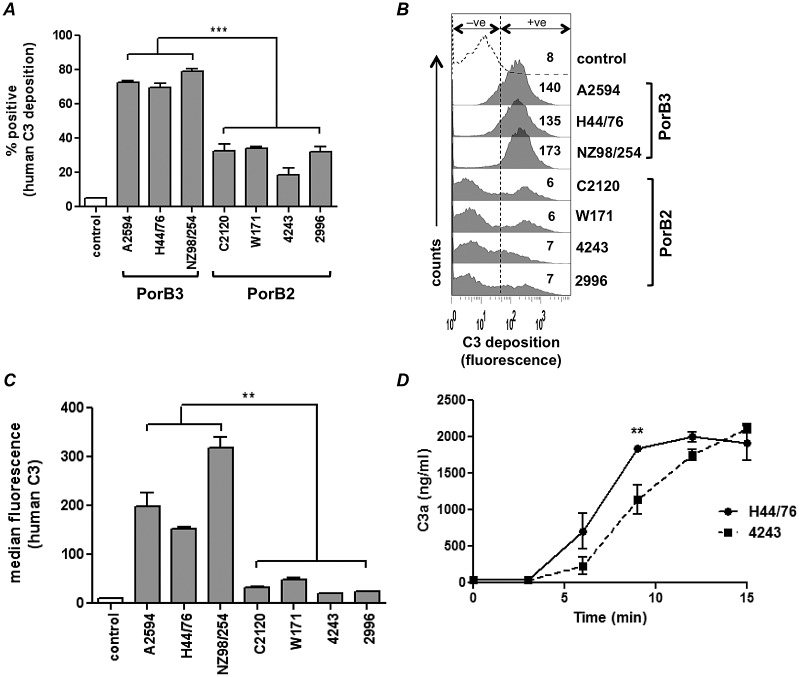
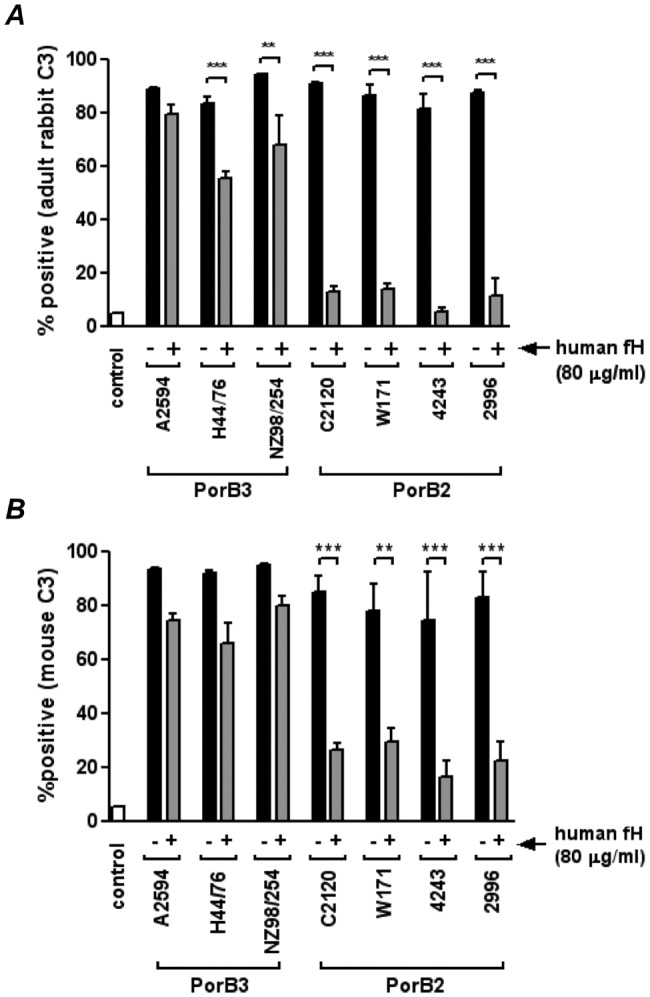
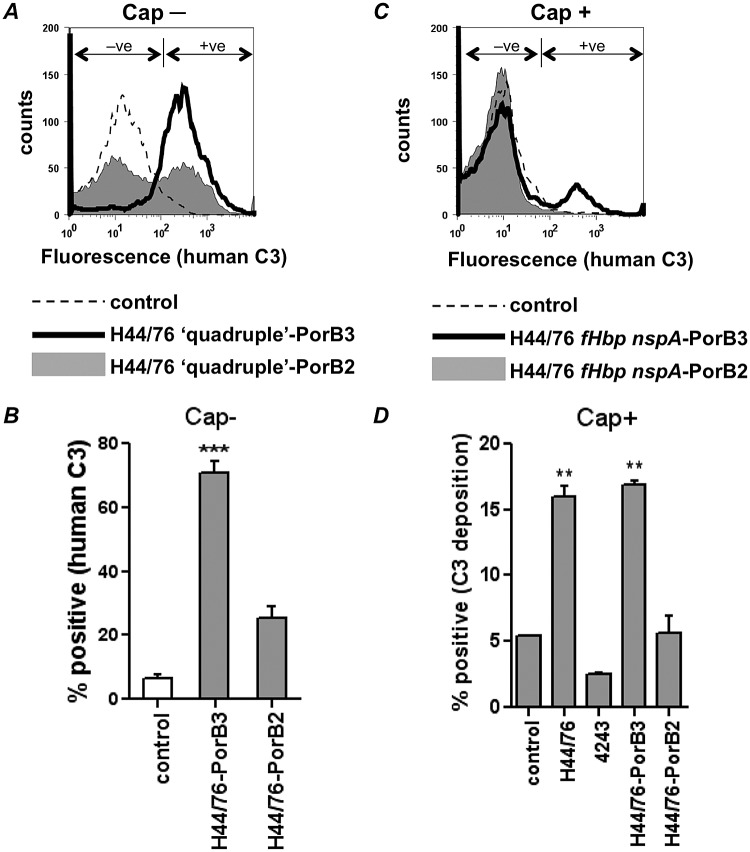
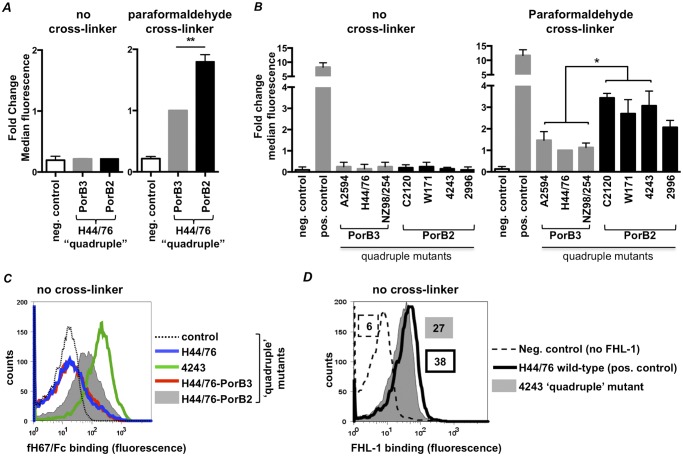
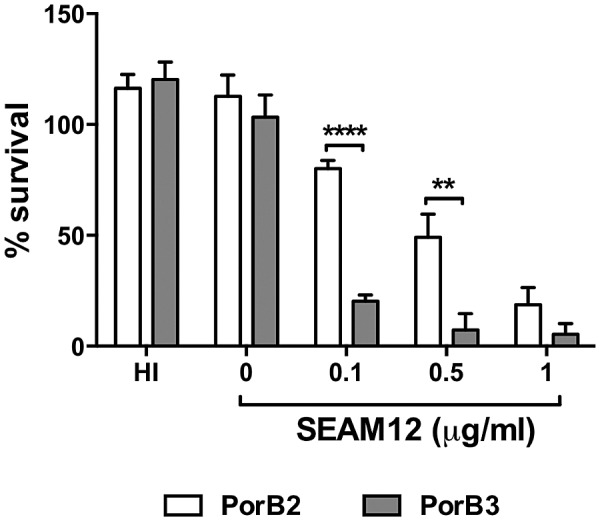
The identification of "factor H binding protein (fHbp)-null" invasive meningococcal isolates and the realization that widespread use of fHbp-based vaccines could herald selection of such strains prompted us to characterize novel mechanisms of alternative pathway (AP) inhibition on meningococci. Of seven strains engineered to lack four known AP-inhibiting molecules, capsular polysaccharide, lipooligosaccharide sialic acid, fHbp, and neisserial surface protein A (quadruple mutants), four strains inhibited human AP-mediated C3 deposition. All four expressed the porin B2 (PorB2) molecule, and three strains belonged to the hypervirulent ST-11 lineage. Consistent with reduced C3 deposition, the rate of C3a generation by a PorB2 isolate was lower than that by a PorB3 strain. Allelic replacement of PorB3 with PorB2, in both encapsulated and unencapsulated strains, confirmed the role of PorB2 in AP inhibition. Expression of PorB2 increased resistance to complement-dependent killing relative to that seen in an isogenic PorB3-expressing strain. Adult rabbit and mouse APs were unimpeded on all mutants, and human fH inhibited nonhuman C3 deposition on PorB2-expressing strains, which provided functional evidence for human fH-dependent AP regulation by PorB2. Low-affinity binding of full-length human fH to quadruple mutants expressing PorB2 was demonstrated. fH-like protein 1 (FHL-1; contains fH domains 1 through 7) and fH domains 6 and 7 fused to IgG Fc bound to one PorB2-expressing quadruple mutant, which suggested that fH domains 6 and 7 may interact with PorB2. These results associate PorB2 expression with serum resistance and presage the appearance of fHbp-null and hypervirulent ST-11 isolates that may evade killing by fHbp-based vaccines.
Importance: The widespread use of antimeningococcal vaccines based on factor H (fH) binding protein (fHbp) is imminent. Meningococci that lack fHbp were recently isolated from persons with invasive disease, and these fHbp-null strains could spawn vaccine failure. Our report provides a molecular basis for an explanation of how fHbp-null strains may evade the host immune system. Meningococci possess several mechanisms to subvert killing by the alternative pathway (AP) of complement, including production of the fHbp and NspA fH binding proteins. Here we show that a meningococcal protein called porin B2 (PorB2) contributes to inhibition of the AP on the bacterial surface. A majority of the "fHbp-null" isolates identified, as well as all members of a "hypervirulent" lineage (called ST-11), express PorB2. Our findings highlight the potential for the emergence of fHbp-negative strains that are able to regulate the AP and may be associated with fHbp vaccine failure.
Figures
Similar articles
-
Binding of complement factor H to PorB3 and NspA enhances resistance of Neisseria meningitidis to anti-factor H binding protein bactericidal activity.Infect Immun. 2015 Apr;83(4):1536-45. doi: 10.1128/IAI.02984-14. Epub 2015 Feb 2. Infect Immun. 2015. PMID: 25644002 Free PMC article.
-
Inhibition of the alternative pathway of nonhuman infant complement by porin B2 contributes to virulence of Neisseria meningitidis in the infant rat model.Infect Immun. 2014 Jun;82(6):2574-84. doi: 10.1128/IAI.01517-14. Epub 2014 Mar 31. Infect Immun. 2014. PMID: 24686052 Free PMC article.
-
The relative roles of factor H binding protein, neisserial surface protein A, and lipooligosaccharide sialylation in regulation of the alternative pathway of complement on meningococci.J Immunol. 2012 May 15;188(10):5063-72. doi: 10.4049/jimmunol.1103748. Epub 2012 Apr 13. J Immunol. 2012. PMID: 22504643 Free PMC article.
-
Does binding of complement factor H to the meningococcal vaccine antigen, factor H binding protein, decrease protective serum antibody responses?Clin Vaccine Immunol. 2013 Aug;20(8):1099-107. doi: 10.1128/CVI.00260-13. Epub 2013 Jun 5. Clin Vaccine Immunol. 2013. PMID: 23740919 Free PMC article. Review.
-
Role of factor H binding protein in Neisseria meningitidis virulence and its potential as a vaccine candidate to broadly protect against meningococcal disease.Microbiol Mol Biol Rev. 2013 Jun;77(2):234-52. doi: 10.1128/MMBR.00056-12. Microbiol Mol Biol Rev. 2013. PMID: 23699256 Free PMC article. Review.
Cited by
-
Binding of complement factor H to PorB3 and NspA enhances resistance of Neisseria meningitidis to anti-factor H binding protein bactericidal activity.Infect Immun. 2015 Apr;83(4):1536-45. doi: 10.1128/IAI.02984-14. Epub 2015 Feb 2. Infect Immun. 2015. PMID: 25644002 Free PMC article.
-
Effect of complement Factor H on anti-FHbp serum bactericidal antibody responses of infant rhesus macaques boosted with a licensed meningococcal serogroup B vaccine.Vaccine. 2015 Dec 16;33(51):7168-7175. doi: 10.1016/j.vaccine.2015.10.135. Epub 2015 Nov 10. Vaccine. 2015. PMID: 26562320 Free PMC article.
-
Hijacking Complement Regulatory Proteins for Bacterial Immune Evasion.Front Microbiol. 2016 Dec 20;7:2004. doi: 10.3389/fmicb.2016.02004. eCollection 2016. Front Microbiol. 2016. PMID: 28066340 Free PMC article. Review.
-
Human IgG1, IgG3, and IgG3 Hinge-Truncated Mutants Show Different Protection Capabilities against Meningococci Depending on the Target Antigen and Epitope Specificity.Clin Vaccine Immunol. 2016 Aug 5;23(8):698-706. doi: 10.1128/CVI.00193-16. Print 2016 Aug. Clin Vaccine Immunol. 2016. PMID: 27307451 Free PMC article.
-
Meningococcal disease and the complement system.Virulence. 2014 Jan 1;5(1):98-126. doi: 10.4161/viru.26515. Epub 2013 Oct 8. Virulence. 2014. PMID: 24104403 Free PMC article. Review.
References
-
- Austen KF, Fearon DT. 1979. A molecular basis of activation of the alternative pathway of human complement. Adv. Exp. Med. Biol. 120B:3–17 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- AI 082263/AI/NIAID NIH HHS/United States
- R37 AI032725/AI/NIAID NIH HHS/United States
- C06 RR 016226/RR/NCRR NIH HHS/United States
- R56 AI032725/AI/NIAID NIH HHS/United States
- R01 AI046464/AI/NIAID NIH HHS/United States
- AI054544/AI/NIAID NIH HHS/United States
- U19 AI084048/AI/NIAID NIH HHS/United States
- AI046464/AI/NIAID NIH HHS/United States
- L40 AI057605/AI/NIAID NIH HHS/United States
- AI084048/AI/NIAID NIH HHS/United States
- R01 AI082263/AI/NIAID NIH HHS/United States
- C06 RR016226/RR/NCRR NIH HHS/United States
- R01 AI032725/AI/NIAID NIH HHS/United States
- R01 AI054544/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
