

The general phosphotransferase system proteins localize to sites of strong negative curvature in bacterial cells
- PMID: 24129255
- PMCID: PMC3812706
- DOI: 10.1128/mBio.00443-13
The general phosphotransferase system proteins localize to sites of strong negative curvature in bacterial cells
Abstract
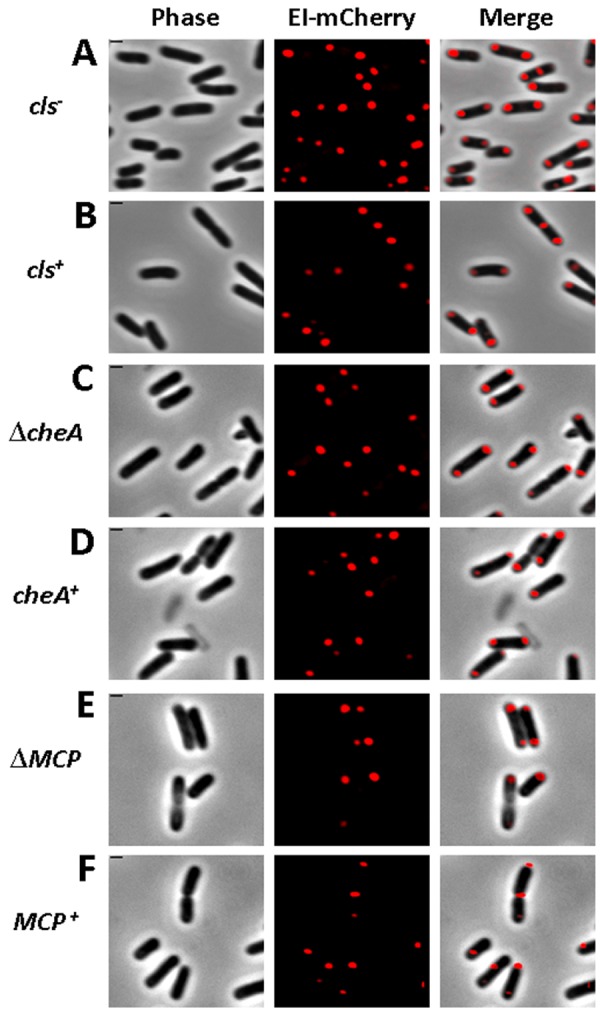
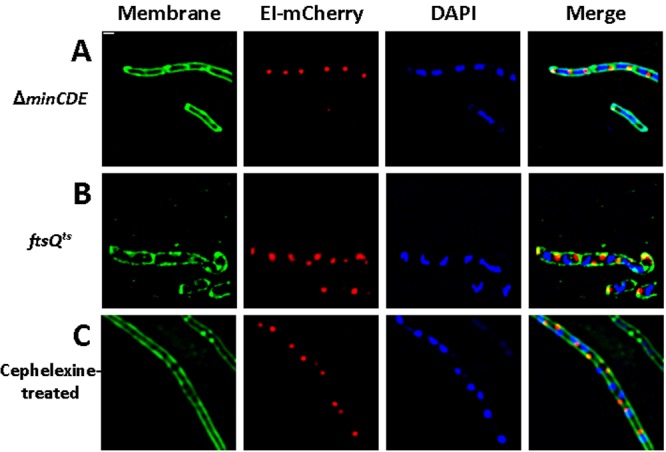
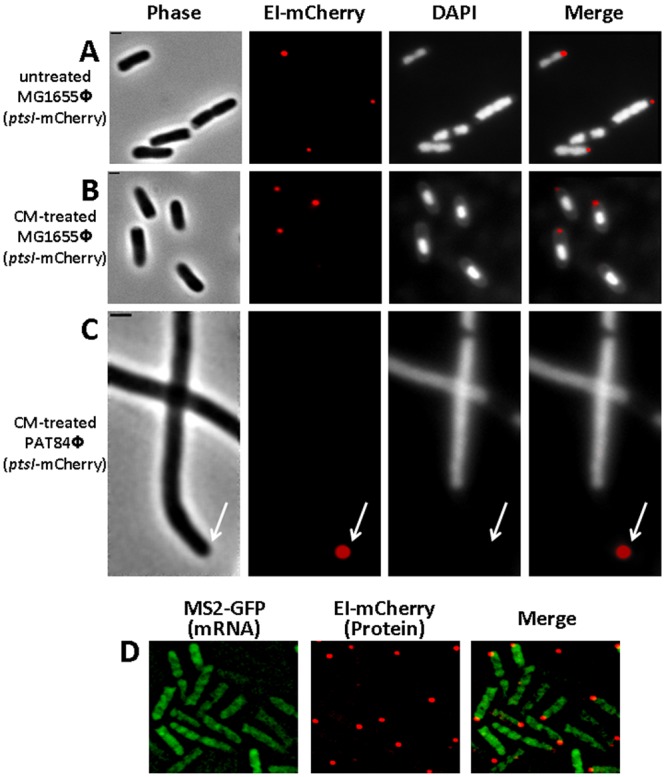
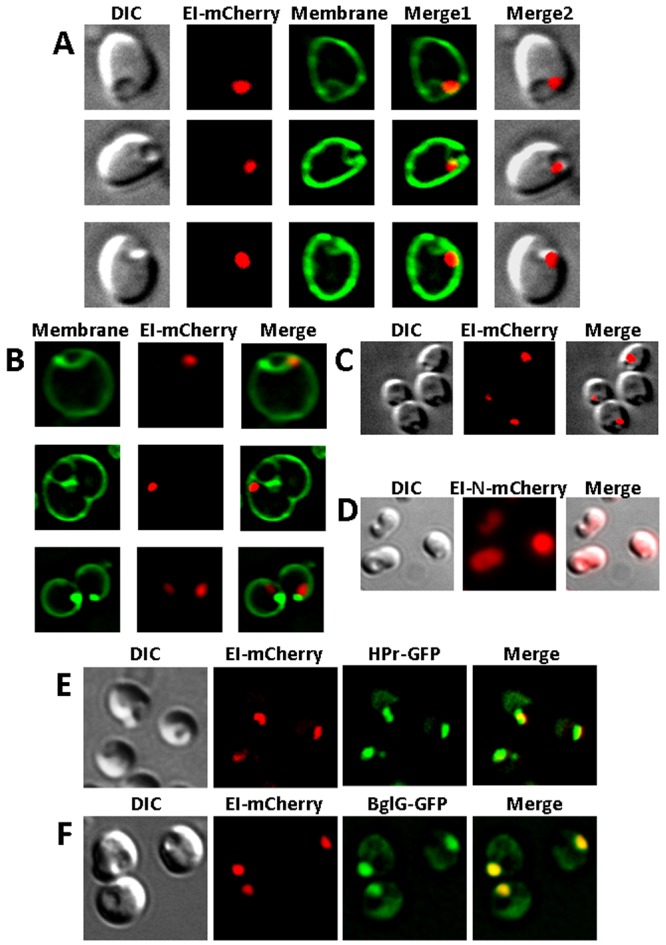
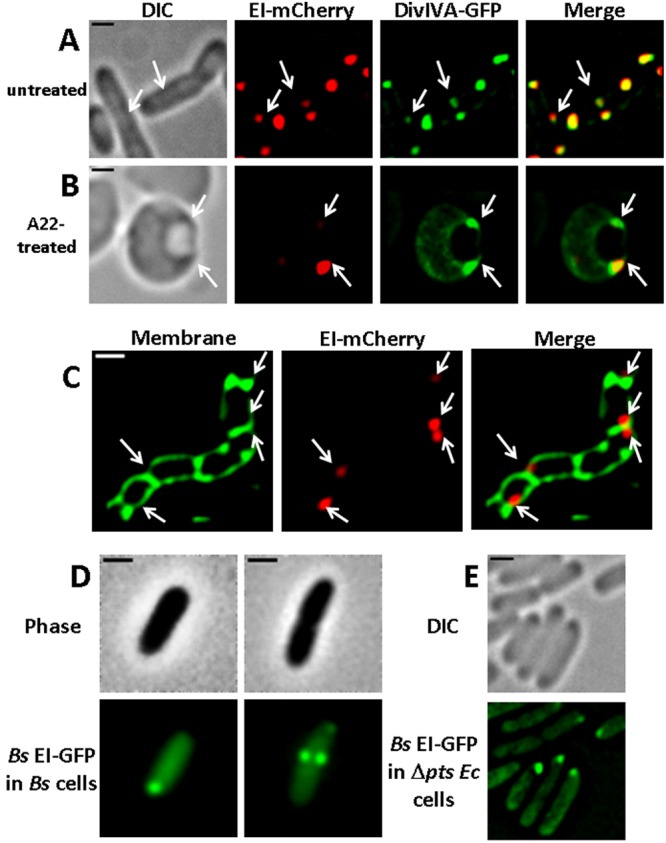
The bacterial cell poles are emerging as subdomains where many cellular activities take place, but the mechanisms for polar localization are just beginning to unravel. The general phosphotransferase system (PTS) proteins, enzyme I (EI) and HPr, which control preferential use of carbon sources in bacteria, were recently shown to localize near the Escherichia coli cell poles. Here, we show that EI localization does not depend on known polar constituents, such as anionic lipids or the chemotaxis receptors, and on the cell division machinery, nor can it be explained by nucleoid occlusion or localized translation. Detection of the general PTS proteins at the budding sites of endocytotic-like membrane invaginations in spherical cells and their colocalization with the negative curvature sensor protein DivIVA suggest that geometric cues underlie localization of the PTS system. Notably, the kinetics of glucose uptake by spherical and rod-shaped E. coli cells are comparable, implying that negatively curved "pole-like" sites support not only the localization but also the proper functioning of the PTS system in cells with different shapes. Consistent with the curvature-mediated localization model, we observed the EI protein from Bacillus subtilis at strongly curved sites in both B. subtilis and E. coli. Taken together, we propose that changes in cell architecture correlate with dynamic survival strategies that localize central metabolic systems like the PTS to subcellular domains where they remain active, thus maintaining cell viability and metabolic alertness.
Importance: Despite their tiny size and the scarcity of membrane-bounded organelles, bacteria are capable of sorting macromolecules to distinct subcellular domains, thus optimizing functionality of vital processes. Understanding the cues that organize bacterial cells should provide novel insights into the complex organization of higher organisms. Previously, we have shown that the general proteins of the phosphotransferase system (PTS) signaling system, which governs utilization of carbon sources in bacteria, localize to the poles of Escherichia coli cells. Here, we show that geometric cues, i.e., strong negative membrane curvature, mediate positioning of the PTS proteins. Furthermore, localization to negatively curved regions seems to support the PTS functionality.
Figures
References
-
- Nevo-Dinur K, Govindarajan S, Amster-Choder O. 2012. Subcellular localization of RNA and proteins in prokaryotes. Trends Genet. 28:314–322 - PubMed
-
- Govindarajan S, Nevo-Dinur K, Amster-Choder O. 2012. Compartmentalization and spatio-temporal organization of macromolecules in bacteria. FEMS Microbiol. Rev. 36:1005–1022 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
