

Ocular phenotype of Fbn2-null mice
- PMID: 24130178
- PMCID: PMC3816615
- DOI: 10.1167/iovs.13-12687
Ocular phenotype of Fbn2-null mice
Abstract
Purpose: Fibrillin-2 (Fbn2) is the dominant fibrillin isoform expressed during development of the mouse eye. To test its role in morphogenesis, we examined the ocular phenotype of Fbn2(-/-) mice.
Methods: Ocular morphology was assessed by confocal microscopy using antibodies against microfibril components.
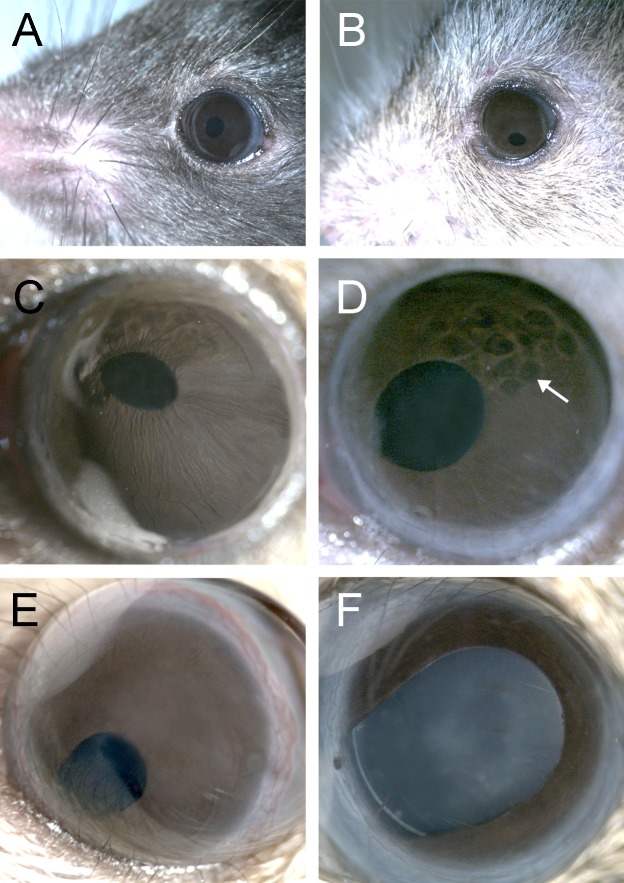
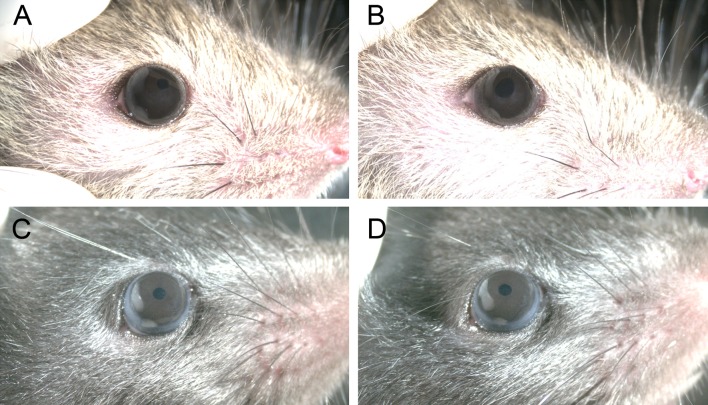
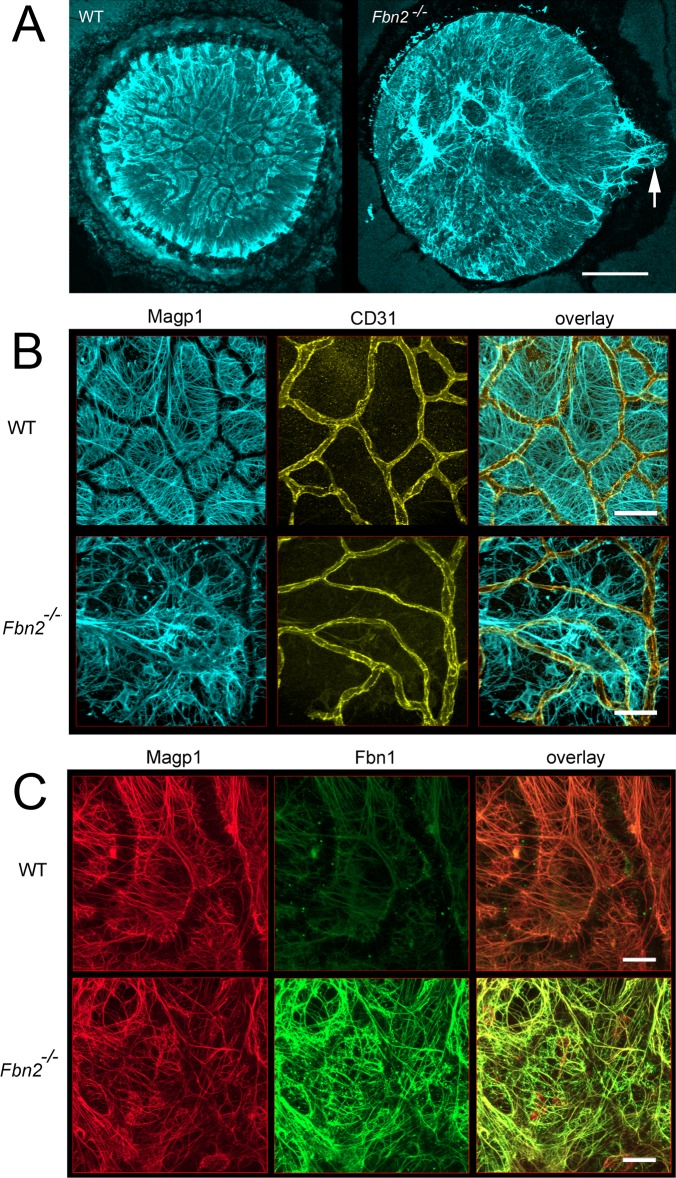
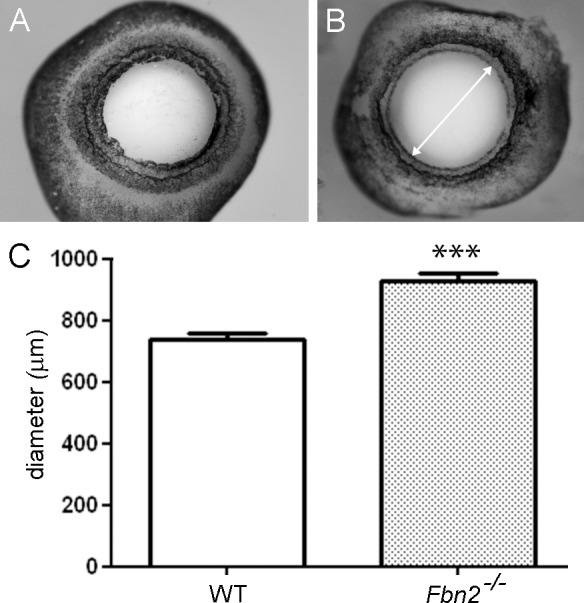
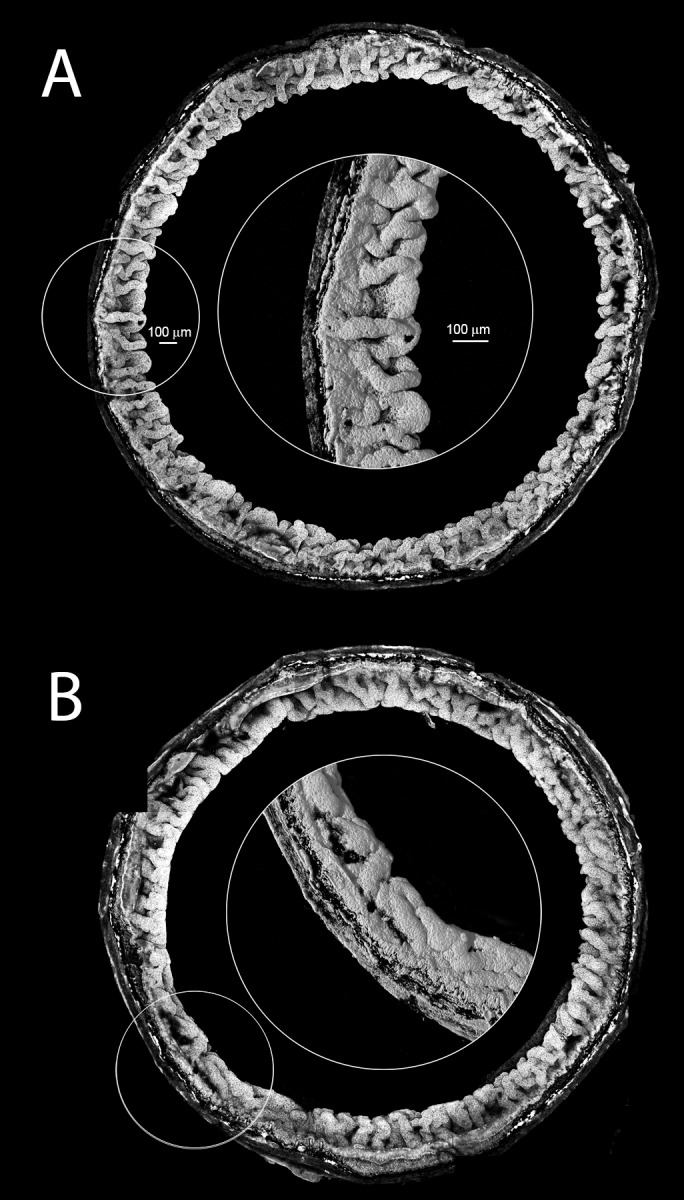
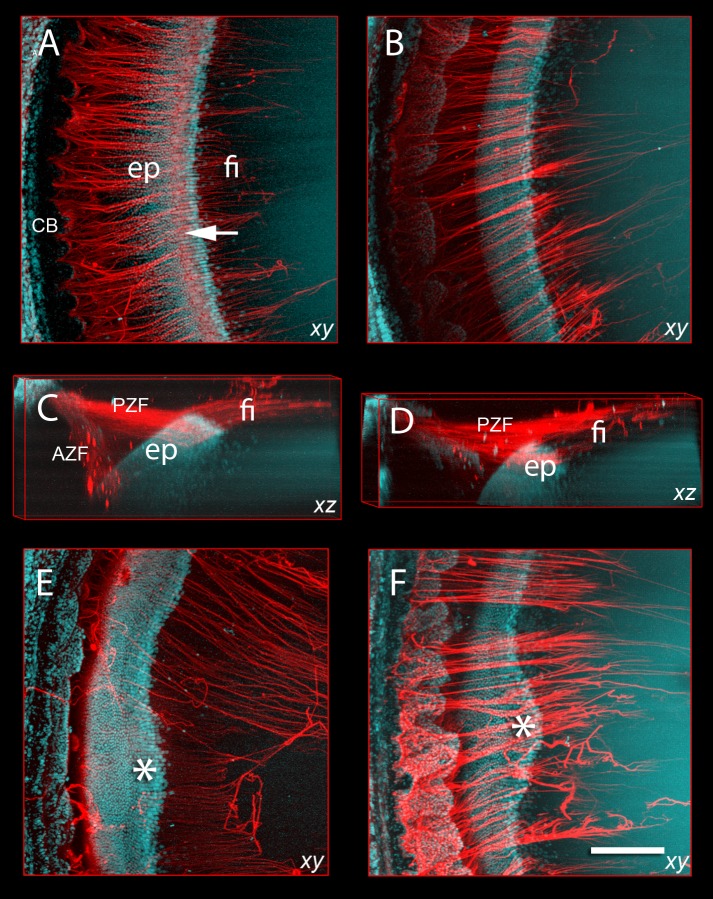
Results: Fbn2(-/-) mice had a high incidence of anterior segment dysgenesis. The iris was the most commonly affected tissue. Complete iridal coloboma was present in 37% of eyes. Dyscoria, corectopia and pseudopolycoria were also common (43% combined incidence). In wild-type (WT) mice, fibrillin-2-rich microfibrils are prominent in the pupillary membrane (PM) during development. In Fbn2-null mice, the absence of Fbn2 was partially compensated for by increased expression of fibrillin-1, although the resulting PM microfibrils were disorganized, compared with WTs. In colobomatous adult Fbn2(-/-) eyes, the PM failed to regress normally, especially beneath the notched region of the iris. Segments of the ciliary body were hypoplastic, and zonular fibers, although relatively plentiful, were unevenly distributed around the lens equator. In regions where the zonular fibers were particularly disturbed, the synchronous differentiation of the underlying lens fiber cells was affected.
Conclusions: Fbn2 has an indispensable role in ocular morphogenesis in mice. The high incidence of iris coloboma in Fbn2-null animals implies a previously unsuspected role in optic fissure closure. The observation that fiber cell differentiation was disturbed in Fbn2(-/-) mice raises the possibility that the attachment of zonular fibers to the lens surface may help specify the equatorial margin of the lens epithelium.
Keywords: ciliary epithelium; ciliary zonule; coloboma; fibrillin; lens; optic fissure; pupillary membrane; tunica vasculosa lentis.
Figures


Similar articles
-
Development, composition, and structural arrangements of the ciliary zonule of the mouse.Invest Ophthalmol Vis Sci. 2013 Apr 1;54(4):2504-15. doi: 10.1167/iovs.13-11619. Invest Ophthalmol Vis Sci. 2013. PMID: 23493297 Free PMC article.
-
Nonselective assembly of fibrillin 1 and fibrillin 2 in the rodent ocular zonule and in cultured cells: implications for Marfan syndrome.Invest Ophthalmol Vis Sci. 2013 Dec 23;54(13):8337-44. doi: 10.1167/iovs.13-13121. Invest Ophthalmol Vis Sci. 2013. PMID: 24265020 Free PMC article.
-
Fibrillins 1 and 2 perform partially overlapping functions during aortic development.J Biol Chem. 2006 Mar 24;281(12):8016-23. doi: 10.1074/jbc.M511599200. Epub 2005 Dec 28. J Biol Chem. 2006. PMID: 16407178 Free PMC article.
-
Zinn's zonule.Prog Retin Eye Res. 2021 May;82:100902. doi: 10.1016/j.preteyeres.2020.100902. Epub 2020 Sep 25. Prog Retin Eye Res. 2021. PMID: 32980533 Free PMC article. Review.
-
Mutations in the human gene for fibrillin-1 (FBN1) in the Marfan syndrome and related disorders.Hum Mol Genet. 1995;4 Spec No:1799-809. doi: 10.1093/hmg/4.suppl_1.1799. Hum Mol Genet. 1995. PMID: 8541880 Review.
Cited by
-
Missense variants of FBN2 associated with congenital arachnodactyly in three Chinese families.Mol Genet Metab Rep. 2024 Sep 9;41:101140. doi: 10.1016/j.ymgmr.2024.101140. eCollection 2024 Dec. Mol Genet Metab Rep. 2024. PMID: 39310807 Free PMC article.
-
Proteomic Analysis of the Bovine and Human Ciliary Zonule.Invest Ophthalmol Vis Sci. 2017 Jan 1;58(1):573-585. doi: 10.1167/iovs.16-20866. Invest Ophthalmol Vis Sci. 2017. PMID: 28125844 Free PMC article.
-
Altered Ocular Fibrillin Microfibril Composition in Mice With a Glaucoma-Causing Mutation of Adamts10.Invest Ophthalmol Vis Sci. 2021 Aug 2;62(10):26. doi: 10.1167/iovs.62.10.26. Invest Ophthalmol Vis Sci. 2021. PMID: 34424262 Free PMC article.
-
Nonproliferative and Proliferative Lesions of the Ratand Mouse Special Sense Organs(Ocular [eye and glands], Olfactory and Otic).J Toxicol Pathol. 2018;31(3 Suppl):97S-214S. doi: 10.1293/tox.31.97S. Epub 2018 Jul 28. J Toxicol Pathol. 2018. PMID: 30158741 Free PMC article. Review. No abstract available.
-
Optimizing Genomic Methods for Mapping and Identification of Candidate Variants in ENU Mutagenesis Screens Using Inbred Mice.G3 (Bethesda). 2018 Feb 2;8(2):401-409. doi: 10.1534/g3.117.300292. G3 (Bethesda). 2018. PMID: 29208648 Free PMC article.
References
-
- Corson GM, Charbonneau NL, Keene DR, Sakai LY. Differential expression of fibrillin-3 adds to microfibril variety in human and avian, but not rodent, connective tissues. Genomics. 2004; 83: 461–472 - PubMed
-
- Jensen SA, Robertson IB, Handford PA. Dissecting the fibrillin microfibril: structural insights into organization and function. Structure. 2012; 20: 215–225 - PubMed
-
- Beals RK, Hecht F. Congenital contractural arachnodactyly. A heritable disorder of connective tissue. J Bone Joint Surg Am. 1971; 53: 987–993 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
