

New world bats harbor diverse influenza A viruses
- PMID: 24130481
- PMCID: PMC3794996
- DOI: 10.1371/journal.ppat.1003657
New world bats harbor diverse influenza A viruses
Abstract
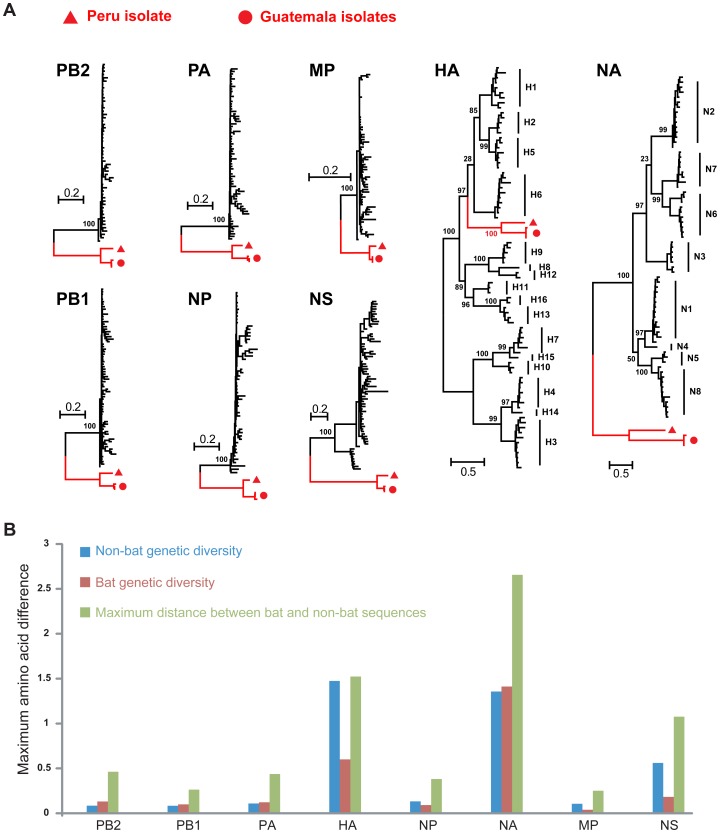
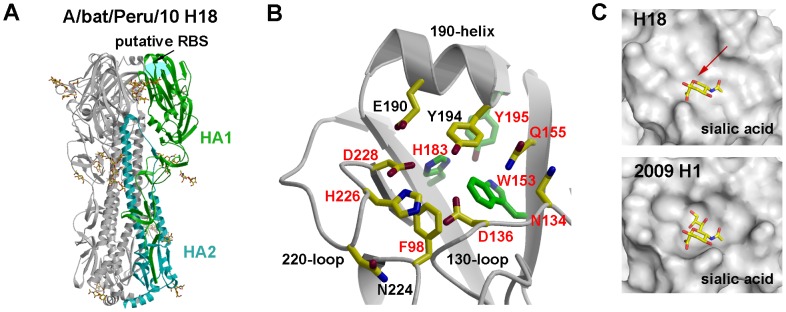
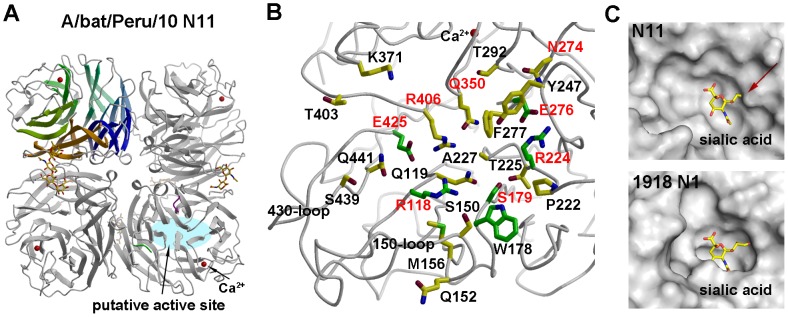
Aquatic birds harbor diverse influenza A viruses and are a major viral reservoir in nature. The recent discovery of influenza viruses of a new H17N10 subtype in Central American fruit bats suggests that other New World species may similarly carry divergent influenza viruses. Using consensus degenerate RT-PCR, we identified a novel influenza A virus, designated as H18N11, in a flat-faced fruit bat (Artibeus planirostris) from Peru. Serologic studies with the recombinant H18 protein indicated that several Peruvian bat species were infected by this virus. Phylogenetic analyses demonstrate that, in some gene segments, New World bats harbor more influenza virus genetic diversity than all other mammalian and avian species combined, indicative of a long-standing host-virus association. Structural and functional analyses of the hemagglutinin and neuraminidase indicate that sialic acid is not a ligand for virus attachment nor a substrate for release, suggesting a unique mode of influenza A virus attachment and activation of membrane fusion for entry into host cells. Taken together, these findings indicate that bats constitute a potentially important and likely ancient reservoir for a diverse pool of influenza viruses.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Li W, Shi Z, Yu M, Ren W, Smith C, et al. (2005) Bats are natural reservoirs of SARS-like coronaviruses. Science 310: 676–679. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
