

The selection of low envelope glycoprotein reactivity to soluble CD4 and cold during simian-human immunodeficiency virus infection of rhesus macaques
- PMID: 24131720
- PMCID: PMC3911730
- DOI: 10.1128/JVI.01558-13
The selection of low envelope glycoprotein reactivity to soluble CD4 and cold during simian-human immunodeficiency virus infection of rhesus macaques
Abstract
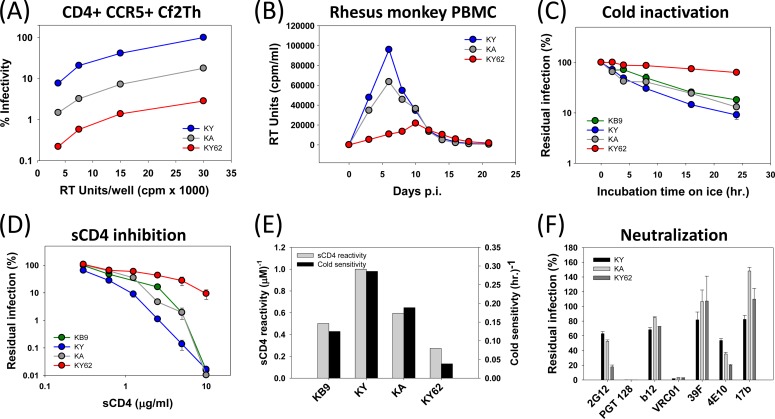
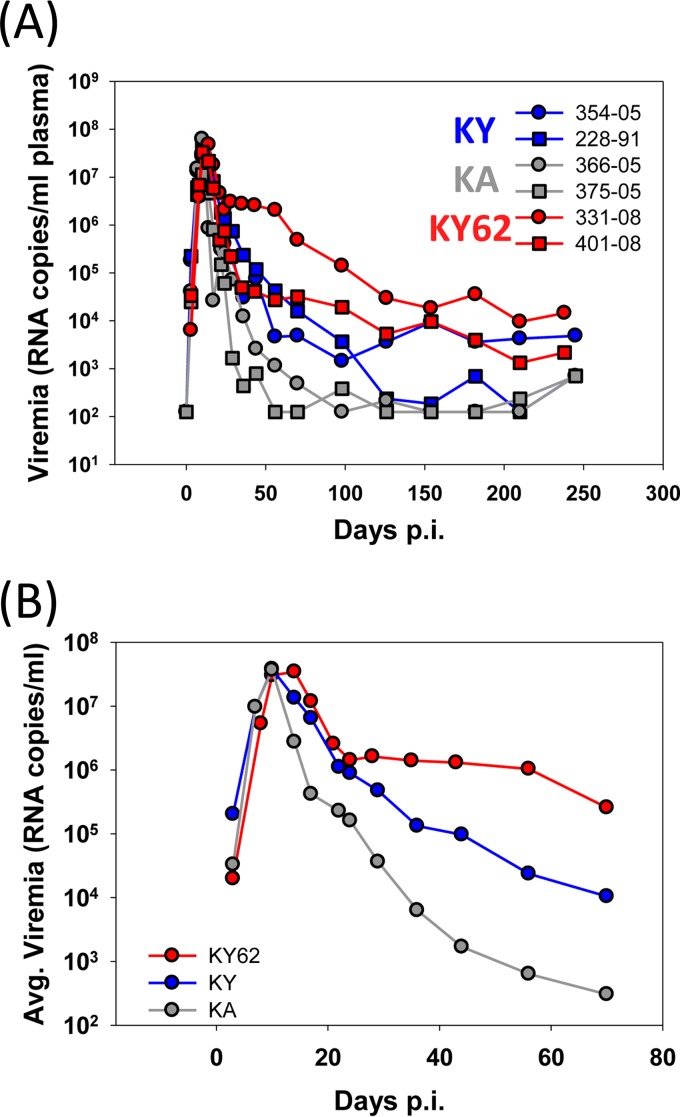
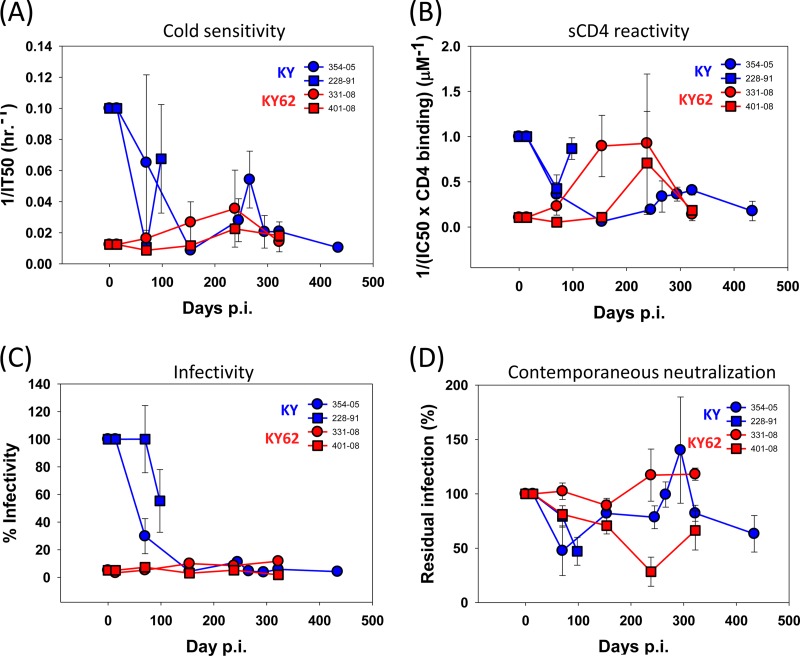
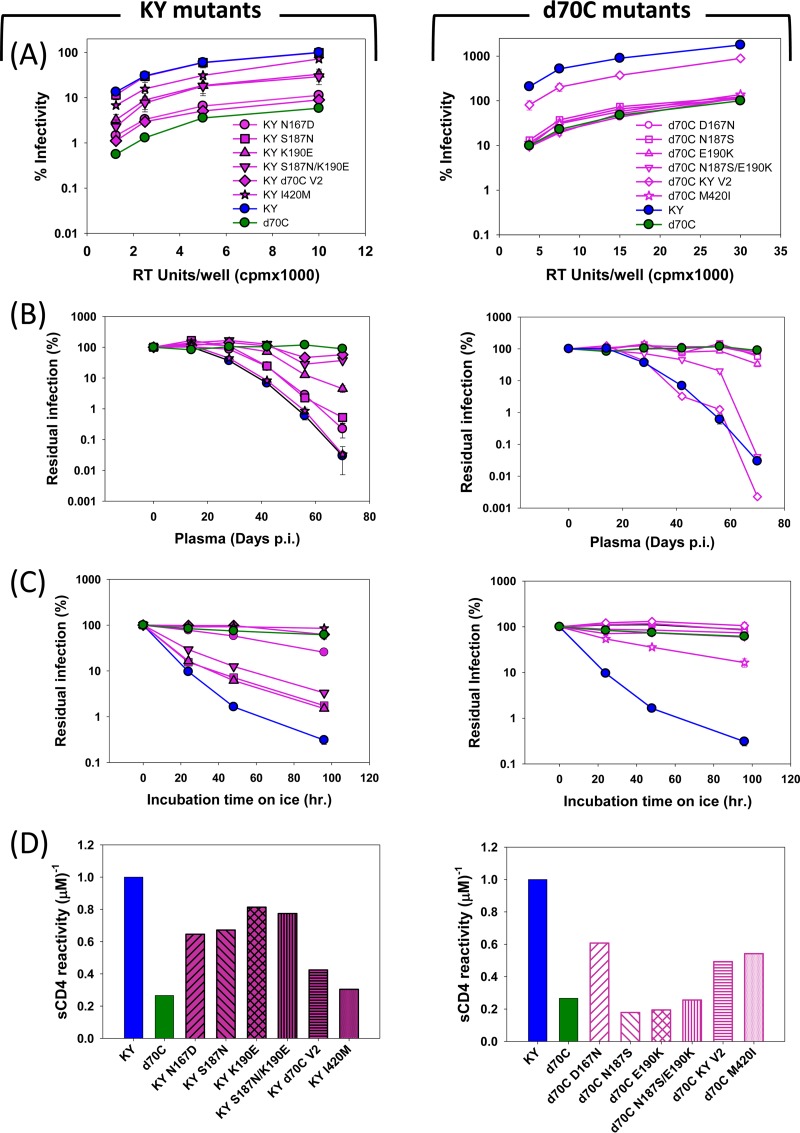
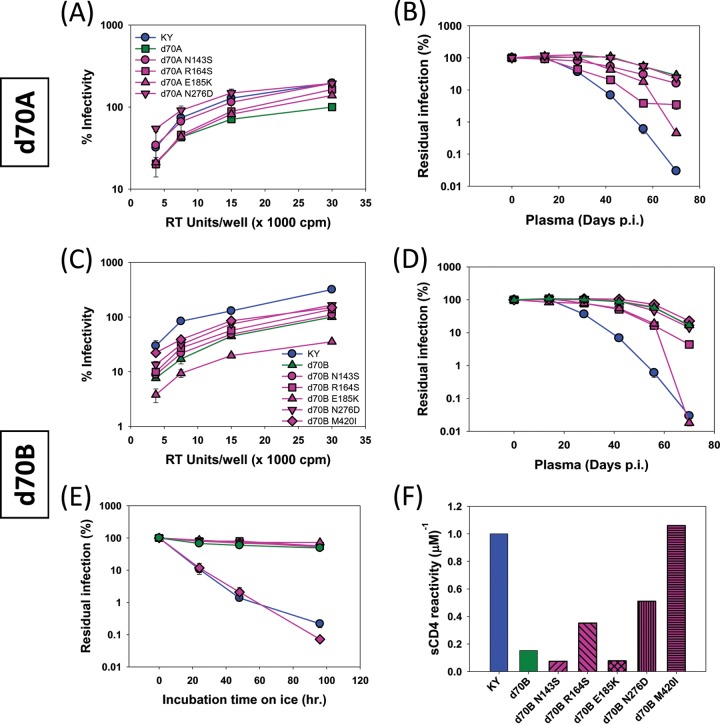
Envelope glycoprotein (Env) reactivity (ER) describes the propensity of human immunodeficiency virus type 1 (HIV-1) Env to change conformation from the metastable unliganded state in response to the binding of ligands (antibodies and soluble CD4 [sCD4]) or incubation in the cold. To investigate Env properties that favor in vivo persistence, we inoculated rhesus macaques with three closely related CCR5-tropic simian-human immunodeficiency viruses (SHIVs) that differ in ER to cold (ERcold) and ER to sCD4 (ERsCD4); these SHIVs were neutralized by antibodies equivalently and thus were similar in ERantibody. All three SHIVs achieved high levels of acute viremia in the monkeys without alteration of their Env sequences, indicating that neither ERcold nor ERsCD4 significantly influences the establishment of infection. Between 14 and 100 days following infection, viruses with high ERcold and ERsCD4 were counterselected. Remarkably, the virus variant with low ERcold and low ERsCD4 did not elicit a neutralizing antibody response against the infecting virus, despite the generation of high levels of anti-Env antibodies in the infected monkeys. All viruses that achieved persistent viremia escaped from any autologous neutralizing antibodies and exhibited low ERcold and low ERsCD4. One set of gp120 changes determined the decrease in ERcold and ERsCD4, and a different set of gp120 changes determined resistance to autologous neutralizing antibodies. Each set of changes contributed to a reduction in Env-mediated entry. During infection of monkeys, any Env replication fitness costs associated with decreases in ERcold and ERsCD4 may be offset by minimizing the elicitation of autologous neutralizing antibodies.
Figures

Similar articles
-
Conformation of HIV-1 Envelope Governs Rhesus CD4 Usage and Simian-Human Immunodeficiency Virus Replication.mBio. 2022 Feb 22;13(1):e0275221. doi: 10.1128/mbio.02752-21. Epub 2022 Jan 11. mBio. 2022. PMID: 35012342 Free PMC article.
-
Rapid Development of gp120-Focused Neutralizing B Cell Responses during Acute Simian Immunodeficiency Virus Infection of African Green Monkeys.J Virol. 2015 Sep;89(18):9485-98. doi: 10.1128/JVI.01564-15. Epub 2015 Jul 8. J Virol. 2015. PMID: 26157116 Free PMC article.
-
Immunization with wild-type or CD4-binding-defective HIV-1 Env trimers reduces viremia equivalently following heterologous challenge with simian-human immunodeficiency virus.J Virol. 2010 Sep;84(18):9086-95. doi: 10.1128/JVI.01015-10. Epub 2010 Jul 7. J Virol. 2010. PMID: 20610729 Free PMC article.
-
Envelope residue 375 substitutions in simian-human immunodeficiency viruses enhance CD4 binding and replication in rhesus macaques.Proc Natl Acad Sci U S A. 2016 Jun 14;113(24):E3413-22. doi: 10.1073/pnas.1606636113. Epub 2016 May 31. Proc Natl Acad Sci U S A. 2016. PMID: 27247400 Free PMC article.
-
Understanding the basis of CD4(+) T-cell depletion in macaques infected by a simian-human immunodeficiency virus.Vaccine. 2002 May 6;20(15):1934-7. doi: 10.1016/s0264-410x(02)00072-5. Vaccine. 2002. PMID: 11983249 Review.
Cited by
-
The Conformational States of the HIV-1 Envelope Glycoproteins.Trends Microbiol. 2020 Aug;28(8):655-667. doi: 10.1016/j.tim.2020.03.007. Epub 2020 May 14. Trends Microbiol. 2020. PMID: 32418859 Free PMC article. Review.
-
Functional and Highly Cross-Linkable HIV-1 Envelope Glycoproteins Enriched in a Pretriggered Conformation.J Virol. 2022 Apr 27;96(8):e0166821. doi: 10.1128/jvi.01668-21. Epub 2022 Mar 28. J Virol. 2022. PMID: 35343783 Free PMC article.
-
The lipid membrane of HIV-1 stabilizes the viral envelope glycoproteins and modulates their sensitivity to antibody neutralization.J Biol Chem. 2020 Jan 10;295(2):348-362. doi: 10.1074/jbc.RA119.009481. Epub 2019 Nov 22. J Biol Chem. 2020. PMID: 31757809 Free PMC article.
-
Regulation of epitope exposure in the gp41 membrane-proximal external region through interactions at the apex of HIV-1 Env.PLoS Pathog. 2022 May 18;18(5):e1010531. doi: 10.1371/journal.ppat.1010531. eCollection 2022 May. PLoS Pathog. 2022. PMID: 35584191 Free PMC article.
-
A broad HIV-1 inhibitor blocks envelope glycoprotein transitions critical for entry.Nat Chem Biol. 2014 Oct;10(10):845-52. doi: 10.1038/nchembio.1623. Epub 2014 Aug 31. Nat Chem Biol. 2014. PMID: 25174000 Free PMC article.
References
-
- Barre-Sinoussi F, Chermann JC, Rey F, Nugeyre MT, Chamaret S, Gruest J, Dauguet C, Axler-Blin C, Vezinet-Brun F, Rouzioux C, Rozenbaum W, Montagnier L. 1983. Isolation of a T-lymphotropic retrovirus from a patient at risk for acquired immune deficiency syndrome (AIDS). Science 220:868–871. 10.1126/science.6189183 - DOI - PubMed
-
- Gao F, Morrison SG, Robertson DL, Thornton CL, Craig S, Karlsson G, Sodroski J, Morgado M, Galvao-Castro B, von Briesen H, Beddows S, Weber J, Sharp PM, Shaw GM, Hahn BH. 1996. Molecular cloning and analysis of functional envelope genes from human immunodeficiency virus type 1 sequence subtypes A through G. The WHO and NIAID networks for HIV isolation and characterization. J. Virol. 70:1651–1667 - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
