

On the importance of cotranscriptional RNA structure formation
- PMID: 24131802
- PMCID: PMC3851714
- DOI: 10.1261/rna.037390.112
On the importance of cotranscriptional RNA structure formation
Abstract
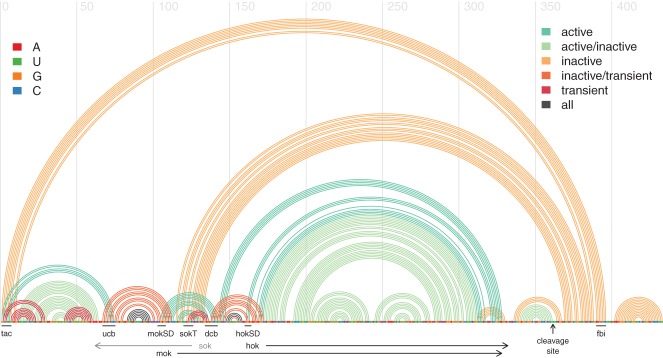
The expression of genes, both coding and noncoding, can be significantly influenced by RNA structural features of their corresponding transcripts. There is by now mounting experimental and some theoretical evidence that structure formation in vivo starts during transcription and that this cotranscriptional folding determines the functional RNA structural features that are being formed. Several decades of research in bioinformatics have resulted in a wide range of computational methods for predicting RNA secondary structures. Almost all state-of-the-art methods in terms of prediction accuracy, however, completely ignore the process of structure formation and focus exclusively on the final RNA structure. This review hopes to bridge this gap. We summarize the existing evidence for cotranscriptional folding and then review the different, currently used strategies for RNA secondary-structure prediction. Finally, we propose a range of ideas on how state-of-the-art methods could be potentially improved by explicitly capturing the process of cotranscriptional structure formation.
Keywords: RNA secondary-structure prediction; RNA structure formation in vivo; cotranscriptional RNA folding.
Figures

References
-
- Alexander R, Barrass J, Dichtl B, Kos M, Obtulowicz T, Robert M, Koper M, Karkusiewicz I, Mariconti L, Tollervey D, et al. 2011. RiboSys, a high-resolution, quantitative approach to measure the in vivo kinetics of pre-mRNA splicing and 3′-end processing in Saccharomyces cerevisiae. RNA 16: 2570–2580 - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous