

Towards greater realism in inclusive fitness models: the case of worker reproduction in insect societies
- PMID: 24132088
- PMCID: PMC3871332
- DOI: 10.1098/rsbl.2013.0334
Towards greater realism in inclusive fitness models: the case of worker reproduction in insect societies
Abstract
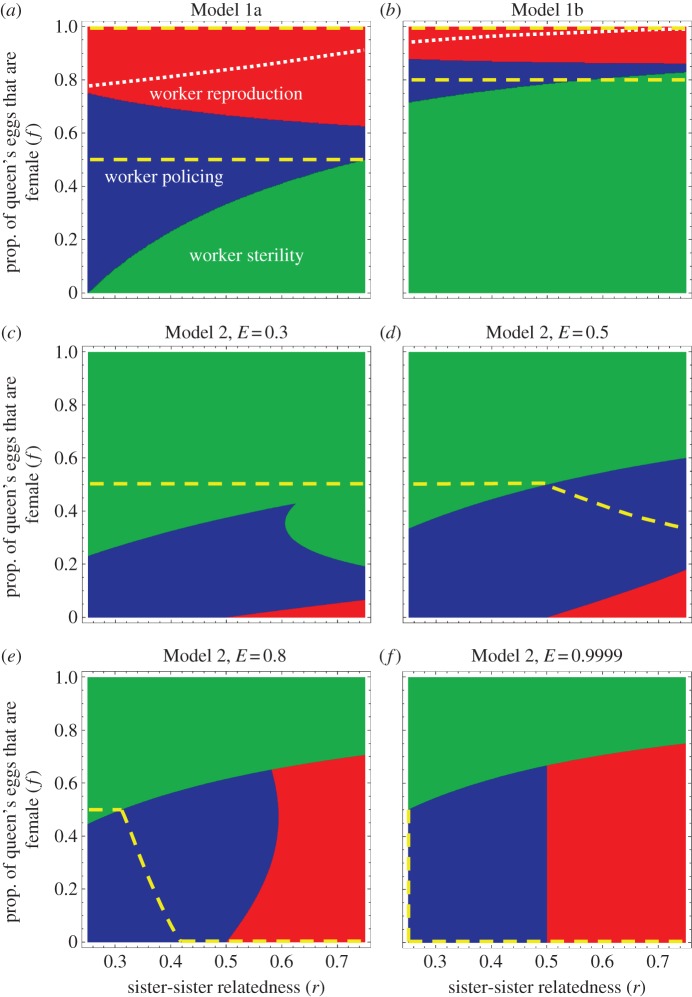
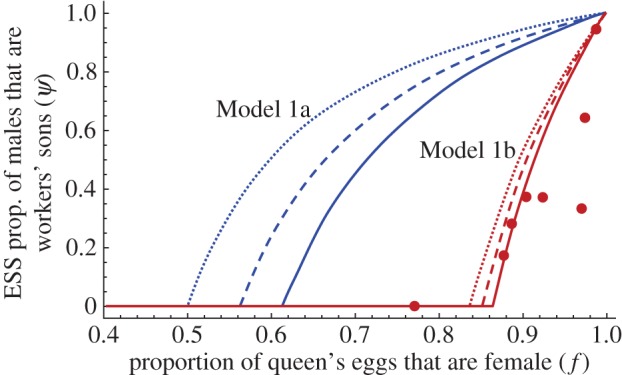
The conflicts over sex allocation and male production in insect societies have long served as an important test bed for Hamilton's theory of inclusive fitness, but have for the most part been considered separately. Here, we develop new coevolutionary models to examine the interaction between these two conflicts and demonstrate that sex ratio and colony productivity costs of worker reproduction can lead to vastly different outcomes even in species that show no variation in their relatedness structure. Empirical data on worker-produced males in eight species of Melipona bees support the predictions from a model that takes into account the demographic details of colony growth and reproduction. Overall, these models contribute significantly to explaining behavioural variation that previous theories could not account for.
Keywords: inclusive fitness; sex allocation; social insects; worker policing; worker reproduction.
Figures
Similar articles
-
Conflict over male parentage in social insects.PLoS Biol. 2004 Sep;2(9):E248. doi: 10.1371/journal.pbio.0020248. Epub 2004 Aug 24. PLoS Biol. 2004. PMID: 15328531 Free PMC article.
-
Conflicts and alliances in insect families.Heredity (Edinb). 2001 May;86(Pt 5):515-21. doi: 10.1046/j.1365-2540.2001.00884.x. Heredity (Edinb). 2001. PMID: 11554967 Review.
-
Mechanisms of social regulation change across colony development in an ant.BMC Evol Biol. 2010 Oct 27;10:328. doi: 10.1186/1471-2148-10-328. BMC Evol Biol. 2010. PMID: 20977775 Free PMC article.
-
Hamilton's rule and the causes of social evolution.Philos Trans R Soc Lond B Biol Sci. 2014 Mar 31;369(1642):20130362. doi: 10.1098/rstb.2013.0362. Print 2014 May 19. Philos Trans R Soc Lond B Biol Sci. 2014. PMID: 24686934 Free PMC article. Review.
-
It's good to be queen: classically eusocial colony structure and low worker fitness in an obligately social sweat bee.Mol Ecol. 2005 Nov;14(13):4123-33. doi: 10.1111/j.1365-294X.2005.02724.x. Mol Ecol. 2005. PMID: 16262863
Cited by
-
Toward greater realism in inclusive fitness models: the case of caste fate conflict in insect societies.Evol Lett. 2024 Jan 11;8(3):387-396. doi: 10.1093/evlett/qrad068. eCollection 2024 Jun. Evol Lett. 2024. PMID: 38818418 Free PMC article.
-
Honeybees possess a structurally diverse and functionally redundant set of queen pheromones.Proc Biol Sci. 2019 Jun 26;286(1905):20190517. doi: 10.1098/rspb.2019.0517. Epub 2019 Jun 19. Proc Biol Sci. 2019. PMID: 31213188 Free PMC article.
-
Cheating and punishment in cooperative animal societies.Philos Trans R Soc Lond B Biol Sci. 2016 Feb 5;371(1687):20150090. doi: 10.1098/rstb.2015.0090. Philos Trans R Soc Lond B Biol Sci. 2016. PMID: 26729930 Free PMC article. Review.
-
Tragedy of the commons in Melipona bees revisited.Biol Lett. 2022 Jan;18(1):20210498. doi: 10.1098/rsbl.2021.0498. Epub 2022 Jan 26. Biol Lett. 2022. PMID: 35078331 Free PMC article.
-
50 Years on: the legacy of William Donald Hamilton.Biol Lett. 2013 Oct 16;9(6):20130792. doi: 10.1098/rsbl.2013.0792. Print 2013. Biol Lett. 2013. PMID: 24132100 Free PMC article. No abstract available.
References
-
- Hamilton WD. 1972. Altruism and related phenomena, mainly in social insects. Annu. Rev. Ecol. Syst. 3, 193–232 (doi:10.1146/annurev.es.03.110172.001205) - DOI
-
- Crozier RH, Pamilo P. 1996. Evolution of social insect colonies. Sex allocation and kin selection. Oxford, UK: Oxford University Press
-
- Ratnieks FLW, Foster KR, Wenseleers T. 2006. Conflict resolution in insect societies. Annu. Rev. Entomol. 51, 581–608 (doi:10.1146/annurev.ento.51.110104.151003) - DOI - PubMed
-
- Boomsma JJ, Grafen A. 1991. Colony level sex ratio selection in the eusocial Hymenoptera. J. Evol. Biol. 4, 383–407 (doi:10.1046/j.1420-9101.1991.4030383.x) - DOI
-
- Bourke AFG. 2005. Genetics, relatedness and social behaviour in insect societies. In Insect evolutionary ecology (eds Fellowes MDE, Holloway GJ, Rolff J.), pp. 1–30 Wallingford, CT: CABI Publishing
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
