

Local Dkk1 crosstalk from breeding ornaments impedes regeneration of injured male zebrafish fins
- PMID: 24135229
- PMCID: PMC3849810
- DOI: 10.1016/j.devcel.2013.08.015
Local Dkk1 crosstalk from breeding ornaments impedes regeneration of injured male zebrafish fins
Abstract
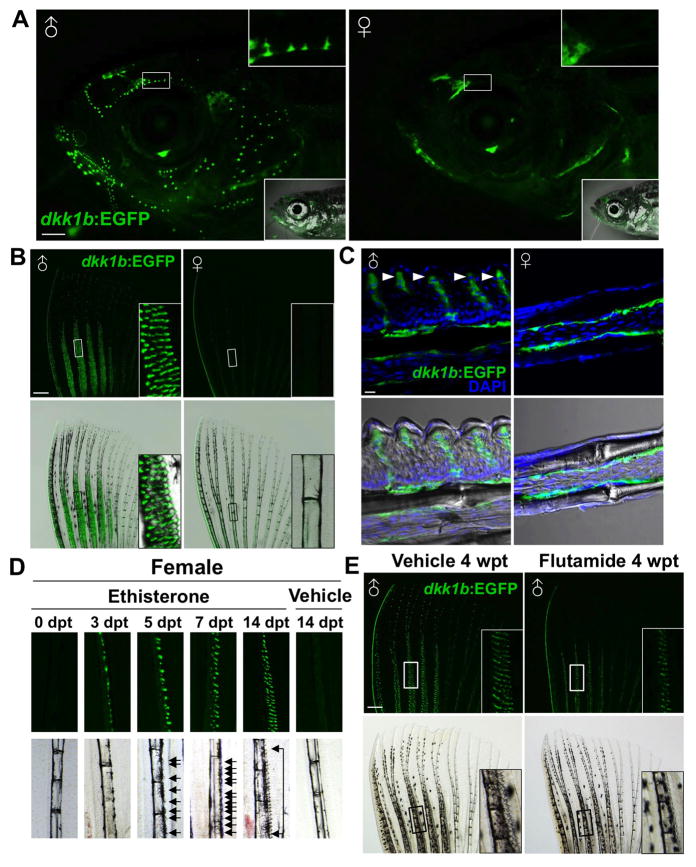
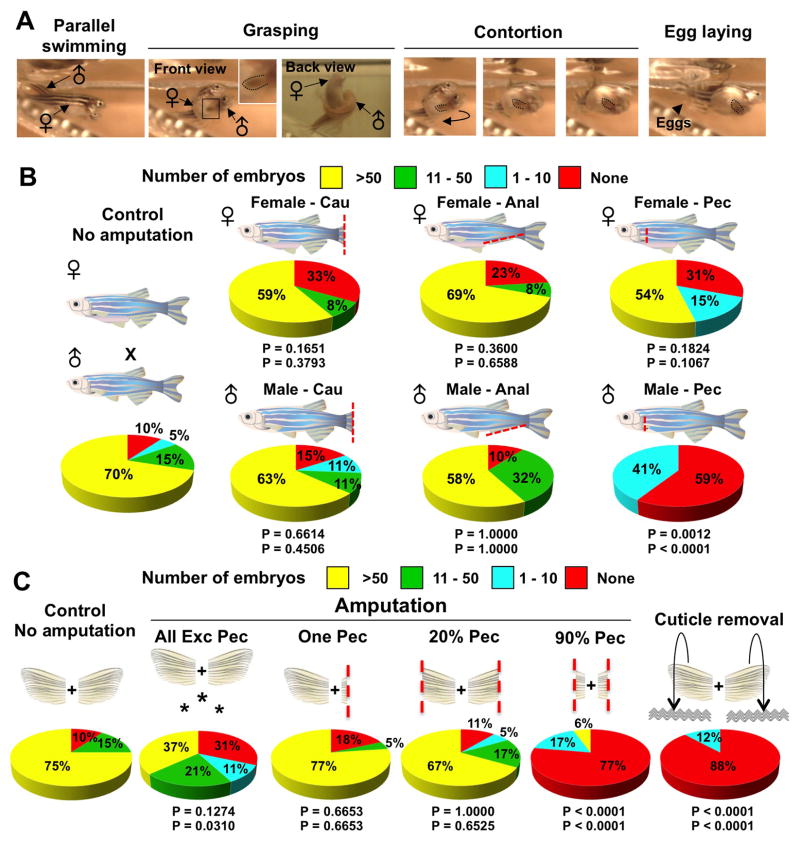
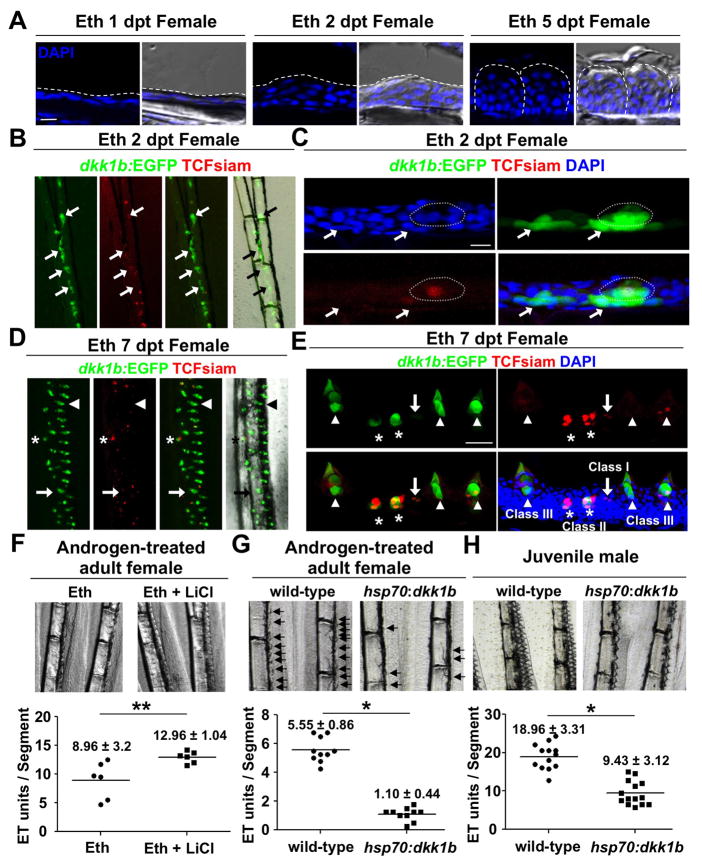
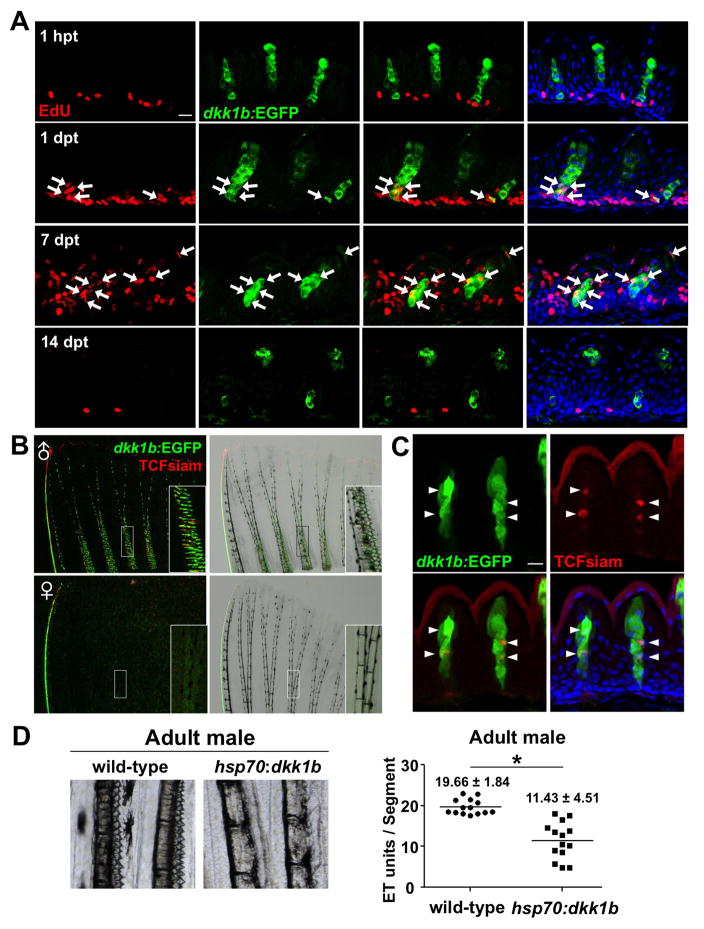
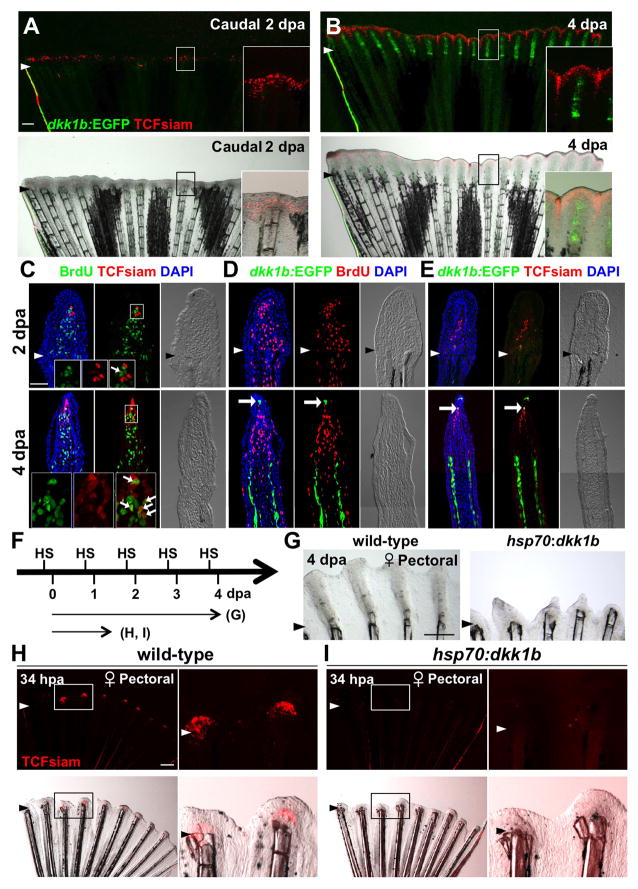
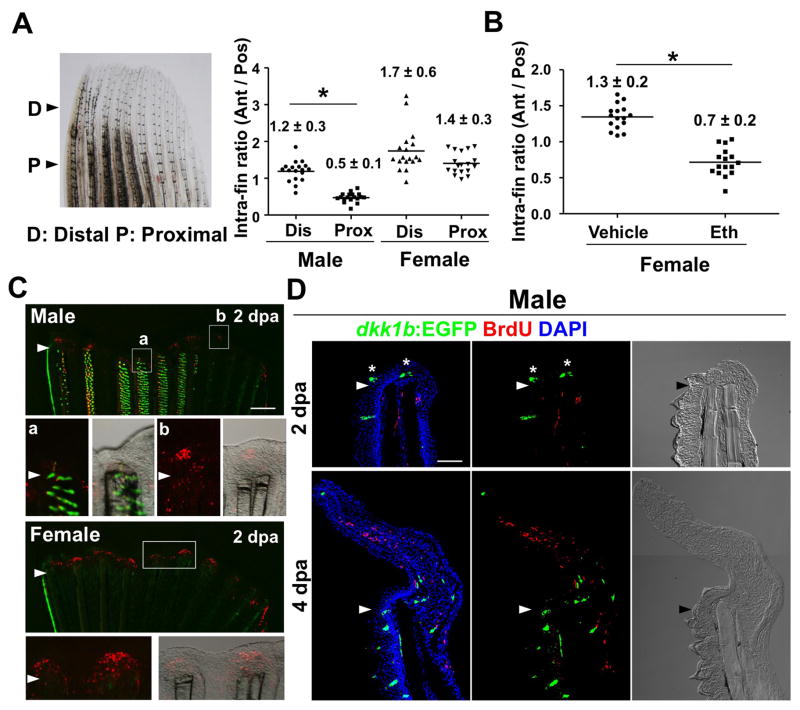
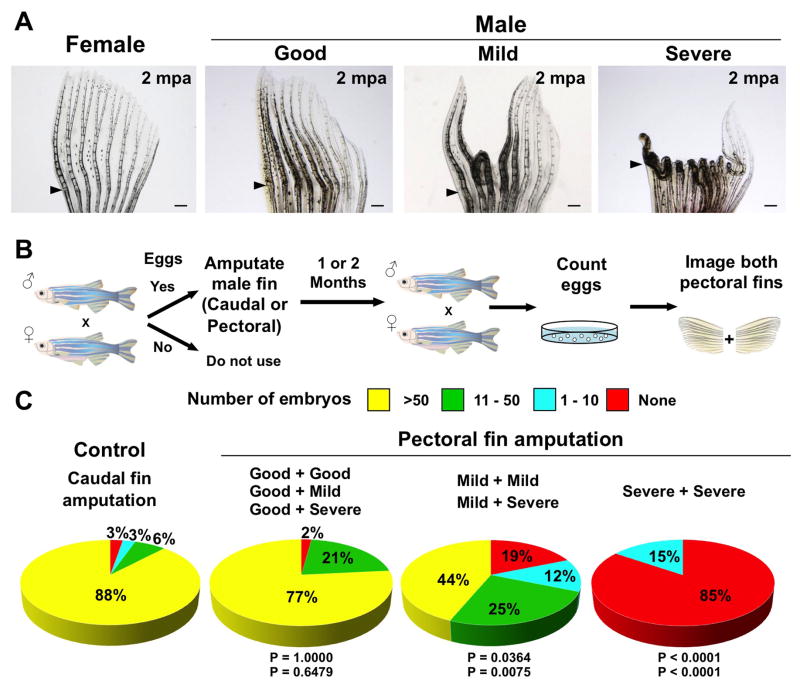
Precise spatiotemporal regulation of signaling activators and inhibitors can help limit developmental crosstalk between neighboring tissues during morphogenesis, homeostasis, and regeneration. Here, we find that the secreted Wnt inhibitor Dkk1b is abundantly produced by dense regions of androgen-regulated epidermal tubercles (ETs) on the surfaces of adult male zebrafish pectoral fins. High-speed videos and amputation experiments reveal that pectoral fins and their ETs are used for male spawning. Formation and vigorous turnover of ETs involve Dkk1b induction and maintenance, whereas Dkk1b is typically restricted from the regeneration blastema after an amputation injury. When amputation occurs through a region containing ETs, a Dkk1b-enriched wound epidermis forms and blastema formation is disrupted, compromising regeneration. Thus, homeostatic signaling by key breeding ornaments can interfere with injury-activated tissue regeneration. Our findings help explain sexually dimorphic fin regeneration in zebrafish and have implications for how regenerative potential might decline as development progresses or during species evolution.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
References
-
- Akimenko MA, Mari-Beffa M, Becerra J, Geraudie J. Old questions, new tools, and some answers to the mystery of fin regeneration. Dev Dyn. 2003;226:190–201. - PubMed
-
- Andl T, Reddy ST, Gaddapara T, Millar SE. WNT signals are required for the initiation of hair follicle development. Dev Cell. 2002;2:643–653. - PubMed
-
- Antos CL, Tanaka EM. Vertebrates that regenerate as models for guiding stem cels. Adv Exp Med Biol. 2010;695:184–214. - PubMed
-
- Bely AE, Nyberg KG. Evolution of animal regeneration: re-emergence of a field. Trends Ecol Evol. 2010;25:161–170. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
