

Crossreactivity of a human autoimmune TCR is dominated by a single TCR loop
- PMID: 24136005
- PMCID: PMC4193804
- DOI: 10.1038/ncomms3623
Crossreactivity of a human autoimmune TCR is dominated by a single TCR loop
Abstract
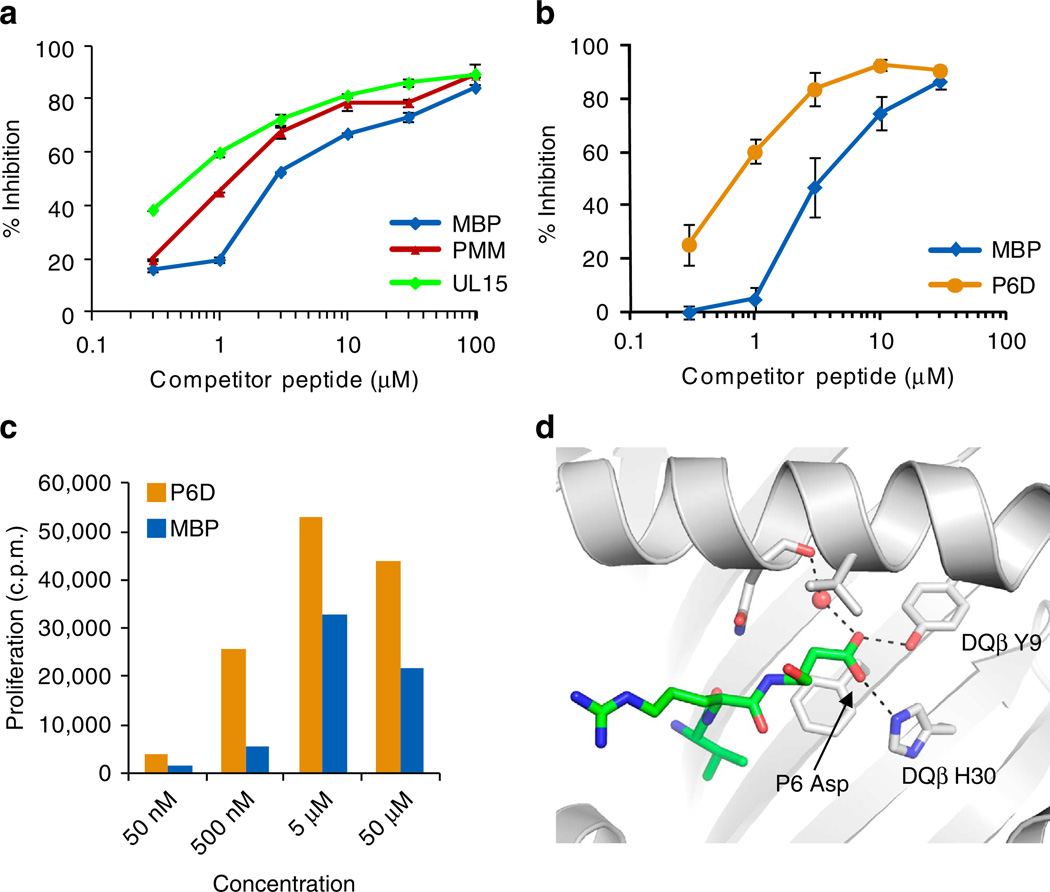
Self-reactive CD4 T cells are thought to have a central role in the pathogenesis of many chronic inflammatory human diseases. Microbial peptides can activate self-reactive T cells, but the structural basis for such crossreactivity is not well understood. The Hy.1B11 T cell receptor (TCR) originates from a patient with multiple sclerosis and recognizes the self-antigen myelin basic protein. Here we report the structural mechanism of TCR crossreactivity with two distinct peptides from human pathogens. The structures show that a single TCR residue (CDR3α F95) makes the majority of contacts with the self-peptide and both microbial peptides (66.7-80.6%) due to a highly tilted TCR-binding topology on the peptide-MHC surface. Further, a neighbouring residue located on the same TCR loop (CDR3α E98) forms an energetically critical interaction with the MHC molecule. These data show how binding by a self-reactive TCR favors crossreactivity between self and microbial antigens.
Figures






Similar articles
-
Unconventional topology of self peptide-major histocompatibility complex binding by a human autoimmune T cell receptor.Nat Immunol. 2005 May;6(5):490-6. doi: 10.1038/ni1187. Epub 2005 Apr 10. Nat Immunol. 2005. PMID: 15821740 Free PMC article.
-
The structural interactions between T cell receptors and MHC-peptide complexes place physical limits on self-nonself discrimination.Curr Top Microbiol Immunol. 2005;296:19-37. doi: 10.1007/3-540-30791-5_2. Curr Top Microbiol Immunol. 2005. PMID: 16329190 Review.
-
Dynamical footprint of cross-reactivity in a human autoimmune T-cell receptor.Sci Rep. 2017 Feb 14;7:42496. doi: 10.1038/srep42496. Sci Rep. 2017. PMID: 28195200 Free PMC article.
-
Positioning of autoimmune TCR-Ob.2F3 and TCR-Ob.3D1 on the MBP85-99/HLA-DR2 complex.Proc Natl Acad Sci U S A. 2008 Oct 7;105(40):15523-8. doi: 10.1073/pnas.0807338105. Epub 2008 Sep 29. Proc Natl Acad Sci U S A. 2008. PMID: 18824684 Free PMC article.
-
Structural basis of molecular mimicry.J Autoimmun. 2001 May;16(3):293-302. doi: 10.1006/jaut.2000.0499. J Autoimmun. 2001. PMID: 11334495 Review.
Cited by
-
Influence of correlated antigen presentation on T-cell negative selection in the thymus.J R Soc Interface. 2018 Nov 7;15(148):20180311. doi: 10.1098/rsif.2018.0311. J R Soc Interface. 2018. PMID: 30404905 Free PMC article.
-
GPU-Accelerated Discovery of Pathogen-Derived Molecular Mimics of a T-Cell Insulin Epitope.Front Immunol. 2020 Feb 28;11:296. doi: 10.3389/fimmu.2020.00296. eCollection 2020. Front Immunol. 2020. PMID: 32184781 Free PMC article.
-
NFM Cross-Reactivity to MOG Does Not Expand a Critical Threshold Level of High-Affinity T Cells Necessary for Onset of Demyelinating Disease.J Immunol. 2017 Oct 15;199(8):2680-2691. doi: 10.4049/jimmunol.1700792. Epub 2017 Sep 8. J Immunol. 2017. PMID: 28887429 Free PMC article.
-
Structural Mechanism Underpinning Cross-reactivity of a CD8+ T-cell Clone That Recognizes a Peptide Derived from Human Telomerase Reverse Transcriptase.J Biol Chem. 2017 Jan 20;292(3):802-813. doi: 10.1074/jbc.M116.741603. Epub 2016 Nov 30. J Biol Chem. 2017. PMID: 27903649 Free PMC article.
-
The good and the bad of T cell cross-reactivity: challenges and opportunities for novel therapeutics in autoimmunity and cancer.Front Immunol. 2023 Jun 19;14:1212546. doi: 10.3389/fimmu.2023.1212546. eCollection 2023. Front Immunol. 2023. PMID: 37409132 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
