

Scanning fluorescence correlation spectroscopy techniques to quantify the kinetics of DNA double strand break repair proteins after γ-irradiation and bleomycin treatment
- PMID: 24137007
- PMCID: PMC3874206
- DOI: 10.1093/nar/gkt908
Scanning fluorescence correlation spectroscopy techniques to quantify the kinetics of DNA double strand break repair proteins after γ-irradiation and bleomycin treatment
Abstract
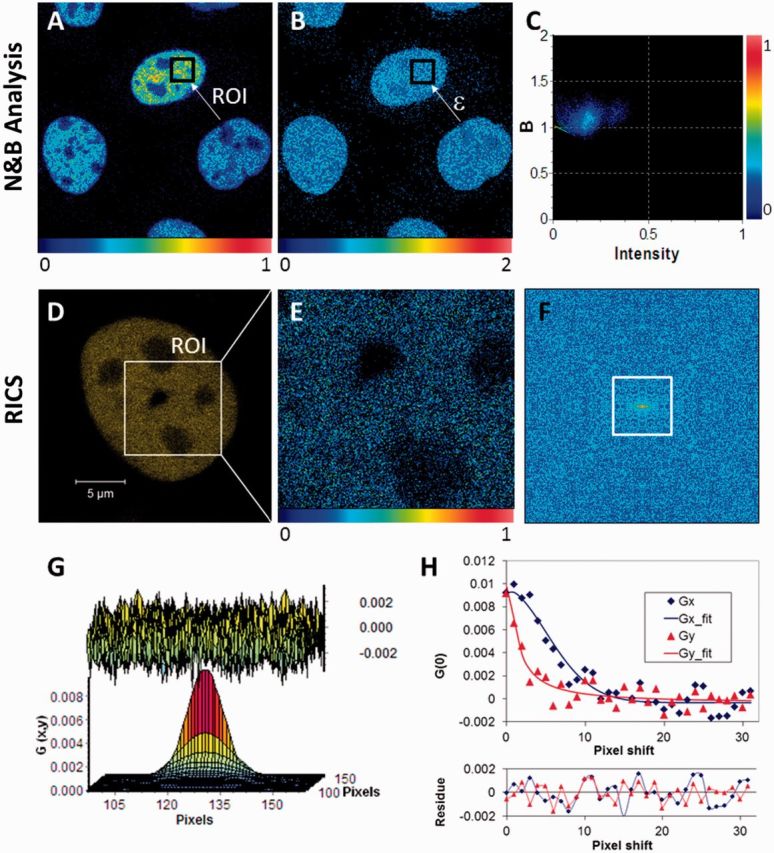
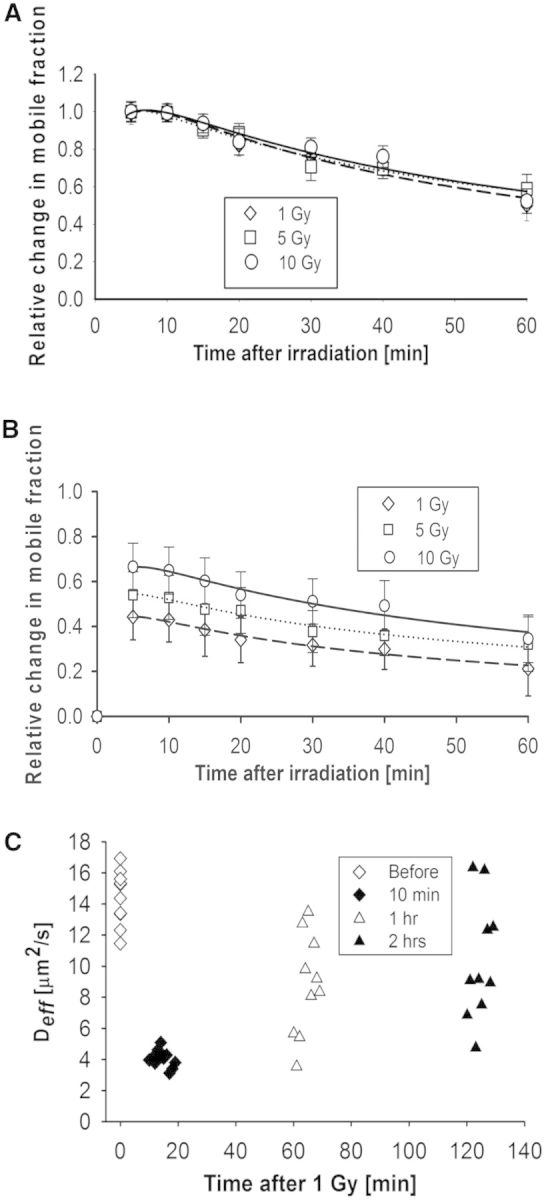
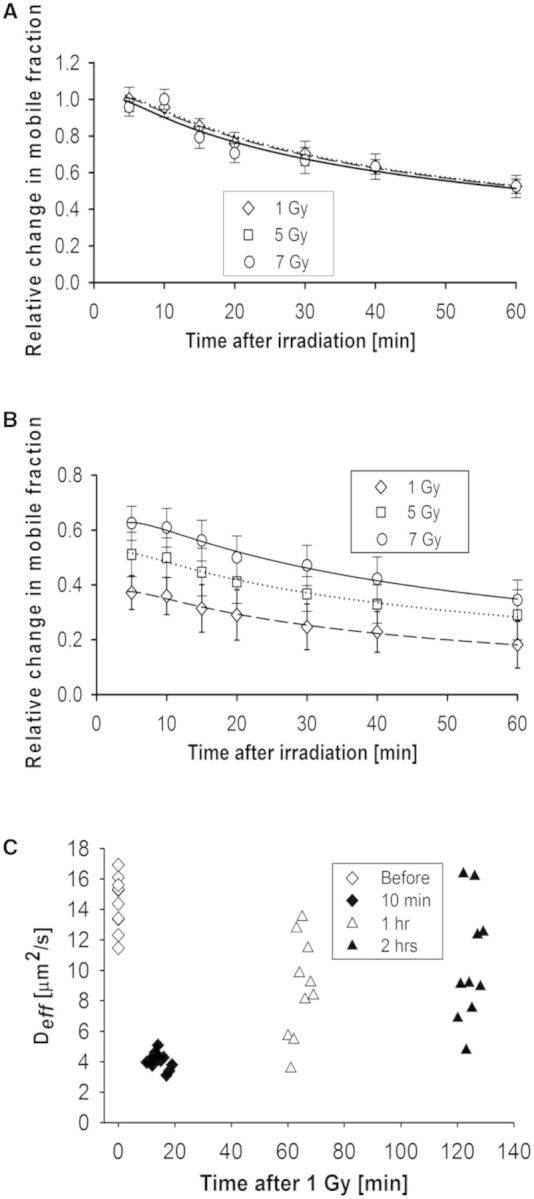
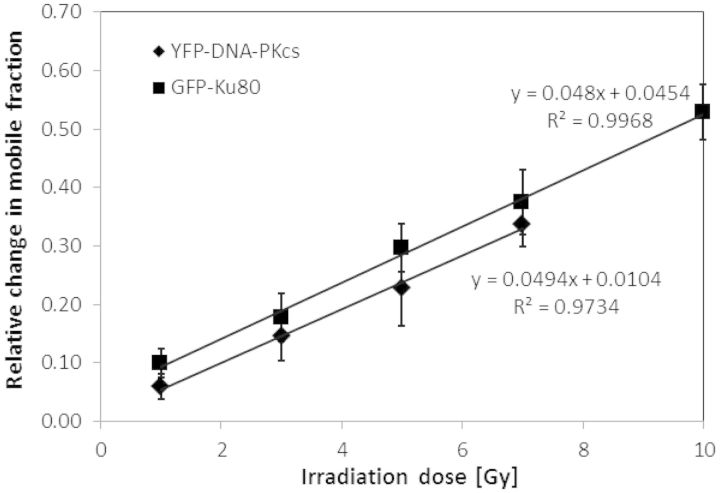
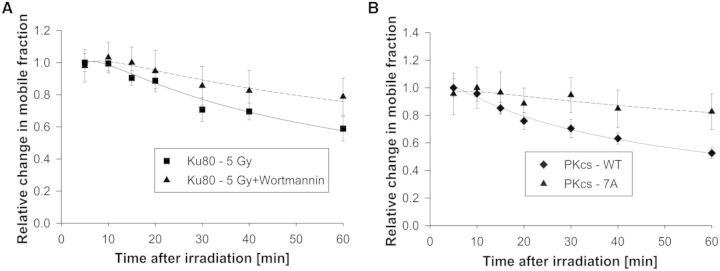
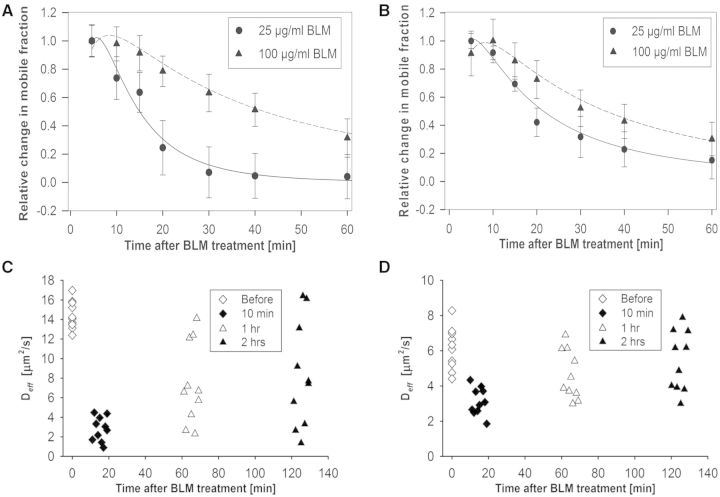
A common feature of DNA repair proteins is their mobilization in response to DNA damage. The ability to visualizing and quantifying the kinetics of proteins localizing/dissociating from DNA double strand breaks (DSBs) via immunofluorescence or live cell fluorescence microscopy have been powerful tools in allowing insight into the DNA damage response, but these tools have some limitations. For example, a number of well-established DSB repair factors, in particular those required for non-homologous end joining (NHEJ), do not form discrete foci in response to DSBs induced by ionizing radiation (IR) or radiomimetic drugs, including bleomycin, in living cells. In this report, we show that time-dependent kinetics of the NHEJ factors Ku80 and DNA-dependent protein kinase catalytic subunits (DNA-PKcs) in response to IR and bleomycin can be quantified by Number and Brightness analysis and Raster-scan Image Correlation Spectroscopy. Fluorescent-tagged Ku80 and DNA-PKcs quickly mobilized in response to IR and bleomycin treatments consistent with prior reports using laser-generated DSBs. The response was linearly dependent on IR dose, and blocking NHEJ enhanced immobilization of both Ku80 and DNA-PKcs after DNA damage. These findings support the idea of using Number and Brightness and Raster-scan Image Correlation Spectroscopy as methods to monitor kinetics of DSB repair proteins in living cells under conditions mimicking radiation and chemotherapy treatments.
Figures
References
-
- Khanna KK, Jackson SP. DNA double-strand breaks: signaling, repair and the cancer connection. Nat. Genet. 2001;27:247–254. - PubMed
-
- Hoeijmakers JH. Genome maintenance mechanisms for preventing cancer. Nature. 2001;411:366–374. - PubMed
-
- Burma S, Chen BP, Chen DJ. Role of non-homologous end joining (NHEJ) in maintaining genomic integrity. DNA Repair. 2006;5:1042–1048. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
